

ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref
Genetic Relations to Ancient North Eurasians (ANE):
Eastern hunter-gatherer (EHG)
Caucasus hunter-gatherer (CHG)
Zagros/Iranian Hunter-Gatherer (IHG)
Iranian Neolithic Farmers (INF)
Anatolian hunter-gatherer (AHG)
Anatolian Neolithic Farmers (ANF)
Early European Farmers (EEF)
Yamnaya/Steppe Herders (WSH)
Villabruna 1 (burial)/Ripari Villabruna rock shelter in northern Italy (14,000 years old)
Satsurblia Cave (burial) in the Country of Georgia (13,000 years old)
Motala (burial) (8,000 years old)
A 23,000-year-old southern Iberian individual links human groups that lived in Western Europe before and after the Last Glacial Maximum
“Following the Bølling/Allerød warming interstadial (14,000 years ago), the Goyet Q2 cluster was replaced by the Villabruna cluster in central Europe, named for its oldest Epigravettian-associated individual from northern Italy, but which also includes most of the Epipalaeolithic- and Mesolithic-associated groups from central and western Europe, all of which are also known as western hunter-gatherers (WHG). In this genetic landscape, Iberian hunter-gatherers (HG) stood out as they retained higher proportions of the Goyet Q2-like ancestry during the Epipalaeolithic and Mesolithic periods and thus are often considered separate.” ref
Human populations underwent range contractions during the Last Glacial Maximum (LGM) which had lasting and dramatic effects on their genetic variation. The genetic ancestry of individuals associated with the post-LGM Magdalenian technocomplex has been interpreted as being derived from groups associated with the pre-LGM Aurignacian. However, both these ancestries differ from that of central European individuals associated with the chronologically intermediate Gravettian. Thus, the genomic transition from pre- to post-LGM remains unclear also in western Europe, where we lack genomic data associated with the intermediate Solutrean, which spans the height of the LGM. Here we present genome-wide data from sites in Andalusia in southern Spain, including from a Solutrean-associated individual from Cueva del Malalmuerzo, directly dated to ~23,000 years ago.” ref
“The Malalmuerzo individual carried genetic ancestry that directly connects earlier Aurignacian-associated individuals with post-LGM Magdalenian-associated ancestry in western Europe. This scenario differs from Italy, where individuals associated with the transition from pre- and post-LGM carry different genetic ancestries. This suggests different dynamics in the proposed southern refugia of Ice Age Europe and posits Iberia as a potential refugium for western European pre-LGM ancestry. Moreover, individuals from Cueva Ardales, which were thought to be of Palaeolithic origin, date younger than expected and, together with individuals from the Andalusian sites Caserones and Aguilillas, fall within the genetic variation of the Neolithic, Chalcolithic, and Bronze Age individuals from southern Iberia.” ref
“Abstract: The concept of grief, the metamorphosis of the deceased into the departed, a subject recreated and rethought by the psyche, is crucial for understanding the significance of the grave and funeral rites. We can divide the funeral rites into three phases: seeing the dead person presented socialized, hiding him to begin the mourning process, and finally metamorphosing him into the deceased. Moreover, these three phases typically require the involvement of several community members, some of whom may be less affected by sorrow — a factor that hinders action — compared to close relatives. Considering these factors, it becomes apparent that grief and, consequently, the tomb are more fundamentally social phenomena than cultural ones. The cultural aspect is an overlay, as beliefs and religions facilitate the mourning process by providing guidelines for conduct and contemplation. An evolutionary perspective on the recognition of death and griefs considers these definitions, cognitive developments during human growth, and the cognitive evolution of hominids. Recognizing another’s death without integrating the concept of one’s mortality could have emerged early in human evolution and been a factor in developing consciousness in a feedback loop. Moreover, the funerary rites and tombs are probably older than is commonly accepted by many researchers to date.” ref
Ancient North Eurasian
“In archaeogenetics, the term Ancient North Eurasian (ANE) is the name given to an ancestral component that represents the lineage of the people of the Mal’ta–Buret’ culture (c. 24,000 years ago) and populations closely related to them, such as the Upper Paleolithic individuals from Afontova Gora in Siberia. Genetic studies also revealed that the ANE are closely related to the remains of the preceding Yana Culture (c. 32,000 years ago), which were dubbed as ‘Ancient North Siberians‘ (ANS). Ancient North Eurasians are predominantly of West Eurasian ancestry (related to European Cro-Magnons and ancient and modern peoples in West Asia) who arrived in Siberia via the “northern route,” but also derive a significant amount of their ancestry (c. 1/3) from an East Eurasian source, having arrived to Siberia via the “southern route.” ref
“Around 20,000 to 25,000 years ago, a branch of Ancient North Eurasian people mixed with Ancient East Asians, which led to the emergence of Ancestral Native American, Ancient Beringian, and Ancient Paleo-Siberian populations. It is unknown exactly where this population admixture took place, and two opposing theories have put forth different migratory scenarios that united the Ancient North Eurasians with ancient East Asian populations. Later, ANE populations migrated westward into Europe and admixed with European Western hunter-gatherer (WHG)-related groups to form the Eastern Hunter-Gatherer (EHG) group, which later admixed with Caucasus hunter-gatherers to form the Western Steppe Herder group, which became widely dispersed across Eurasia during the Bronze Age.” ref
“ANE ancestry has spread throughout Eurasia and the Americas in various migrations since the Upper Paleolithic, and more than half of the world’s population today derives between 5 and 42% of their genomes from the Ancient North Eurasians. Significant ANE ancestry can be found in Native Americans, as well as in Europe, South Asia, Central Asia, and Siberia. It has been suggested that their mythology may have featured narratives shared by both Indo-European and some Native American cultures, such as the existence of a metaphysical world tree and a fable in which a dog guards the path to the afterlife.” ref
Eastern hunter-gatherer
“In archaeogenetics, eastern hunter-gatherer (EHG), sometimes east European hunter-gatherer or eastern European hunter-gatherer, is a distinct ancestral component that represents Mesolithic hunter-gatherers of Eastern Europe. The eastern hunter-gatherer genetic profile is mainly derived from Ancient North Eurasian (ANE) ancestry, which was introduced from Siberia, with a secondary and smaller admixture of European western hunter-gatherers (WHG). Still, the relationship between the ANE and EHG ancestral components is not yet well understood due to lack of samples that could bridge the spatiotemporal gap. During the Mesolithic, the EHGs inhabited an area stretching from the Baltic Sea to the Urals and downwards to the Pontic–Caspian steppe. Along with Scandinavian hunter-gatherers (SHG) and western hunter-gatherers (WHG), the EHGs constituted one of the three main genetic groups in the postglacial period of early Holocene Europe.” ref
“The border between WHGs and EHGs ran roughly from the lower Danube, northward along the western forests of the Dnieper towards the western Baltic Sea. During the Neolithic and early Eneolithic, likely during the 4th millennium BC EHGs on the Pontic–Caspian steppe mixed with Caucasus hunter-gatherers (CHGs) with the resulting population, almost half-EHG and half-CHG, forming the genetic cluster known as Western Steppe Herder (WSH). WSH populations closely related to the people of the Yamnaya culture are supposed to have embarked on a massive migration leading to the spread of Indo-European languages throughout large parts of Eurasia.” ref
Caucasus hunter-gatherer
“Caucasus hunter-gatherer (CHG), also called Satsurblia cluster, is an anatomically modern human genetic lineage, first identified in a 2015 study, based on the population genetics of several modern Western Eurasian (European, Caucasian, and Near Eastern) populations. It represents an ancestry maximized in some Upper Paleolithic and Mesolithic hunter-gatherer groups in the Caucasus. These groups are also very closely related to Mesolithic hunter-gatherers and Neolithic farmers in the Iranian Plateau, who are sometimes included within the CHG group. Ancestry that is closely related to CHG-Iranian Neolithic farmers is also known from further east, including from the Bactria–Margiana Archaeological Complex and the Harappan/Indus Valley Civilisation. Caucasus hunter-gatherers and Eastern hunter-gatherers are ancestral in roughly equal proportions to the Western Steppe Herders (WSH), who were widely spread across Europe and Asia beginning during the Chalcolithic.” ref
“The CHG lineage is suggested to have diverged from the ancestor of Western Hunter-Gatherers (WHGs) probably during the Last Glacial Maximum (sometime between 45,000 to 26,000 years ago). They further separated from the Anatolian hunter-gatherer (AHG) lineage later, suggested to around 25,000 years ago during the late LGM period. The Caucasus hunter-gatherers managed to survive in isolation since the late LGM period as a distinct population, and display high genetic affinities to Mesolithic and Neolithic populations on the Iranian plateau, such as Neolithic specimens found in Ganj Dareh. The CHG display higher genetic affinities to European and Anatolian groups than Iranian hunter-gatherers do, suggesting a possible cline and geneflow into the CHG and less into Mesolithic and Neolithic Iranian groups.” ref
“Lazaridis et. al (2016) models the CHG as a mixture of Neolithic Iranians, Western Hunter-Gatherers, and Eastern Hunter-Gatherers. In addition, the CHG cluster with early Iranian farmers, who significantly do not share alleles with early Levantine farmers. An alternative model without the need of significant amounts of ANE ancestry has been presented by Vallini et al. (2024), suggesting that the initial Iranian hunter-gatherer-like population, which is basal to the CHG formed primarily from a deep Ancient West Eurasian lineage (‘WEC2’, c. 72%), and from varying degrees of Ancient East Eurasian (c. 10%) and Basal Eurasian (c. 18%) components. The Ancient West Eurasian component associated with Iranian hunter-gatherers (WEC2) is inferred to have diverged from the West Eurasian Core lineage (represented by Kostenki-14; WEC), with the WEC2 component staying in the region of the Iranian Plateau, while the proper WEC component expanded into Europe.” ref
“Irving-Pease et. al (2024) models CHG as being derived from an Out of Africa population that split into basal Northern Europeans and West Asians. The latter was where CHG originated from. At the beginning of the Neolithic, at c. 8000 BC, they were probably distributed across western Iran and the Caucasus, and people similar to northern Caucasus and Iranian plateau hunter-gatherers arrived before 6000 BC in Pakistan and north-west India. A roughly equal merger between the CHG and Eastern Hunter-Gatherers in the Pontic–Caspian steppe resulted in the formation of the Western Steppe Herders (WSHs). The WSHs formed the Yamnaya culture and subsequently expanded massively throughout Europe during the Late Neolithic and Early Bronze Age c. 3000—2000 BCE. Caucasus hunter-gatherer/Iranian-like ancestry, was first reported as maximized in hunter-gatherers from the South Caucasus and early herders/farmers in northwestern Iran, particularly the Zagros, hence the label “CHG/Iranian.” ref
Epigravettian
“The Epigravettian (Greek: epi “above, on top of,” and Gravettian ) was one of the last archaeological industries and cultures of the European Upper Paleolithic . It emerged after the Last Glacial Maximum around ~21,000 years ago or 19,050 BCE. It succeeds the Gravettian culture in Italy. Initially named Tardigravettian (Late Gravettian) in 1964 by Georges Laplace in reference to several lithic industries found in Italy, it was later renamed in order to better emphasize its independent character. Three subphases, the Early Epigravettian (20,000 to 16,000 years ago), the Evolved Epigravettian (16,000 to 14,000 years ago), and the Final Epigravettian (14,000 to 8,000 years ago) have been established, which were further subdivided and reclassified. In this sense, the Epigravettian is simply the Gravettian after ~21,000 years ago, when the Solutrean had replaced the Gravettian in most of France and Spain. Epigravettian Venus like artifacts, maybe 24,000 – 18,000 years ago. An Epigravettian ceramic figurine of a horse or deer, Vela Spila, Croatia, is dated to 15,400-14,600 years ago.” ref
“Several Epigravettian cultural centers have developed contemporaneously after 22,000 years years ago in Europe. These range across southern, central, and most of eastern Europe, including southwestern France, Italy, Southeast Europe , the Caucasus, Ukraine, and Western Russia to the banks of the Volga River. Its lithic complex was first documented at numerous sites in Italy. Great geographical and local variability of the facies is present, however all sites are characterized by the predominance of microliths, such as backed blades, backed points, and bladelets with retouched end. The Epigravettian is the last stage of the Upper Paleolithic succeeded by Mesolithic cultures after 10,000 years ago. In a genetic study published in Nature in May 2016, the remains of an Epigravettian male from Ripari Villabruna in Italy were examined.” ref
“He carried the paternal haplogroup R1b1 and the maternal haplogroup U5b . An Epigravettian from the Satsurblia Cave in Georgia , who was examined in a previous study, has been found to be carrying the paternal haplogroup J1 and the maternal haplogroup K3. Analysis of Epigravettian producing individuals in Italy indicates that they not closely related to earlier Gravettian-producing populations of the peninsula, and instead belong to the Villabruna genetic cluster . This group is more closely related to ancient and modern peoples in the Middle East and the Caucasus than earlier European Cro-Magnons . Epigravettian peoples belonging to the Western Hunter Gatherer genetic cluster expanded across Western Europe at the end of the Pleistocene, largely replacing the producers of the Magdalenian culture that previously dominated the region.” ref
“Magdalenian cultures (Pre-R1b Y-DNA Europe) are later cultures of the Upper Paleolithic and Mesolithic in western Europe. They date from around 17,000 to 12,000 years ago. It is named after the type site of La Madeleine, a rock shelter located in the Vézère valley, commune of Tursac, in France’s Dordogne department. The Magdalenian is associated with reindeer hunters, although Magdalenian sites contain extensive evidence for the hunting of red deer, horses, and other large mammals present in Europe toward the end of the last glacial period. The culture was geographically widespread, and later Magdalenian sites stretched from Portugal in the west to Poland in the east, and as far north as France, the Channel Islands, England, and Wales. Besides La Madeleine, the chief stations of the Magdalenian are Les Eyzies, Laugerie-Basse, and Gorges d’Enfer in the Dordogne; Grotte du Placard in Charente and others in south-west France.” ref
“Magdalenian peoples produced a wide variety of art, including figurines and cave paintings. Evidence has been found suggesting that Magdalenian peoples regularly engaged in (probably ritualistic) cannibalism along with producing skull cups. Genetic studies indicate that the Magdalenian peoples were largely descended from earlier Western European Cro-Magnon groups like the Gravettians that were present in Western Europe over 30,000 years ago prior to the Last Glacial Maximum, who had retreated to southwestern Europe during the LGM. Madgalenian peoples were largely replaced and, in some areas, absorbed by Epigravettian producing groups of Villabruna/Western Hunter Gatherer ancestry at the end of the Pleistocene.” ref
“The genes of seven Magdalenians, the El Miron Cluster in Iberia, have shown a close relationship to a population who had lived in Northern Europe some 20,000 years previously. The analyses suggested that 70-80% of the ancestry of these individuals was from the population represented by Goyet Q116-1, associated with the Aurignacian culture of about 35,000 BP, from the Goyet Caves in modern Belgium. It has also been found that Magdalenians are also closely related to western Gravettians who inhabited France and Spain prior to the Last Glacial Maximum. The 15,000-year-old GoyetQ2 individual from Goyet Caves is often used as a proxy for Magdalenian ancestry.” ref
“Analysis of genomes of GoyetQ2-related Magdalenians suggest that, like earlier Cro-Magnon groups, they probably had a relatively dark skin tone compared to modern Europeans. A 2023 study proposed that relative to earlier Western European Cro-Magnon-related groups like Goyet Q116-1-related Aurignacian and the Western Gravettian-associated Fournol cluster, the Goyet-Q2-related Magdalenians appear to have carried significant (~30% ancestry) from the Villabruna cluster (thought to be of southeastern European origin, and sharing affinities to West Asian peoples not found in earlier European hunter-gatherers) associated with the Epigravettian.” ref
“The three samples of Y-DNA included two samples of haplogroup I and one sample of HIJK. All samples of mtDNA belonged to U, including five samples of U8b and one sample of U5b. Around 14-12,000 years ago, the Western Hunter-Gatherer cluster (which predominantly descended from the Villabruna cluster, with possible ancestry related to the Goyet-Q2 cluster), expanded northwards across the Alps, largely replacing the Goyet-Q2 cluster associated Magdalenian groups in Western Europe. In France and Spain, significant GoyetQ2-related ancestry persisted into the Mesolithic and Neolithic, with some Neolithic individuals in France and Spain largely of Early European Farmer descent showing significant GoyetQ2 ancestry.” ref
Western hunter-gatherer
“In archaeogenetics, western hunter-gatherer (WHG, also known as west European hunter-gatherer, western European hunter-gatherer or Oberkassel cluster) (c. 15,000~5,000 years ago) is a distinct ancestral component of modern Europeans, representing descent from a population of Mesolithic hunter-gatherers who scattered over western, southern and central Europe, from the British Isles in the west to the Carpathians in the east, following the retreat of the ice sheet of the Last Glacial Maximum. It is closely associated and sometimes considered synonymous with the concept of the Villabruna cluster, named after Ripari Villabruna cave in Italy, known from the terminal Pleistocene of Europe, which is largely ancestral to later WHG populations.” ref
“WHGs share a closer genetic relationship to ancient and modern peoples in the Middle East and the Caucasus than earlier European hunter-gatherers. Their precise relationships to other groups are somewhat obscure, with their origin likely somewhere in southeast Europe or West Asia. They had expanded into the Italian and Iberian Peninsulas by approximately 19,000 years ago, and subsequently expanded across Western Europe at the end of the Pleistocene around 14-12,000 years ago, largely replacing the Magdalenians who previously dominated the region. The Magdalenians largely descended from earlier Western European Cro-Magnon groups that had arrived in the region over 30,000 years ago, prior to the Last Glacial Maximum.” ref
“WHGs constituted one of the main genetic groups in the postglacial period of early Holocene Europe, along with eastern hunter-gatherers (EHG) in Eastern Europe. The border between WHGs and EHGs ran roughly from the lower Danube, northward along the western forests of the Dnieper towards the western Baltic Sea. EHGs primarily consisted of a mixture of WHG-related and Ancient North Eurasian (ANE) ancestry. Scandinavia was inhabited by Scandinavian hunter-gatherers (SHGs), which were a mixture between WHG and EHG. In the Iberian Peninsula, early Holocene hunter-gathers consisted of a mixture of WHG and Magdalenian Cro-Magnon (GoyetQ2) ancestry.” ref
“Once the main population throughout Europe, the WHGs were largely replaced by successive expansions of Early European Farmers (EEFs) of Anatolian origin during the early Neolithic, who generally carried a minor amount of WHG ancestry due to admixture with WHG groups during their European expansion. Among modern-day populations, WHG ancestry is most common among populations of the eastern Baltic region. Western hunter-gatherers (WHG) are recognised as a distinct ancestral component contributing to the ancestry of most modern Europeans. Most Europeans can be modeled as a mixture of WHG, EEF, and WSH from the Pontic–Caspian steppe. WHGs also contributed ancestry to other ancient groups such as Early European Farmers (EEF), who were, however, mostly of Anatolian descent. With the Neolithic expansion, EEF came to dominate the gene pool in most parts of Europe, although WHG ancestry had a resurgence in Western Europe from the Early Neolithic to the Middle Neolithic.” ref
“WHGs represent a major population shift within Europe at the end of the Ice Age, probably a population expansion into continental Europe, from Southeastern European or West Asian refugia. It is thought that their ancestors separated from eastern Eurasians around 40,000 years ago, and from Ancient North Eurasians (ANE) prior to 24,000 years ago (the estimated age date of the Mal’ta boy). This date was subsequently put further back in time by the findings of the Yana Rhinoceros Horn Site to around 38,000 years ago, shortly after the divergence of West-Eurasian and East-Eurasian lineages. Vallini et al. 2022 argues that the dispersal and split patterns of West Eurasian lineages was not earlier than c. 38,000 years ago, with older Initial Upper Paleolithic European specimens, such as those found in the Zlaty Kun, Peștera cu Oase and Bacho Kiro caves, being unrelated to Western hunter-gatherers but closer to Ancient East Eurasians or basal to both.” ref
“The relationships of the WHG/Villabruna cluster to other Paleolithic human groups in Europe and West Asia are obscure and subject to conflicting intepretations. A 2022 study proposed that the WHG/Villabruna population genetically diverged from hunter-gatherers in the Middle East and the Caucasus around 26,000 years ago, during the Last Glacial Maximum. WHG genomes display higher affinity for ancient and modern Middle Eastern populations when compared against earlier Paleolithic Europeans such as Gravettians. The affinity for ancient Middle Eastern populations in Europe increased after the Last Glacial Maximum, correlating with the expansion of WHG (Villabruna or Oberkassel) ancestry. There is also evidence for bi-directional geneflow between WHG and Middle Eastern populations as early as 15,000 years ago.” ref
“WHG associated remains belonged primarily to the human Y-chromosome haplogroups I-M170 with a lower frequency of C-F3393 (specifically the clade C-V20/C1a2), which has been found commonly among earlier Paleolithic European remains such as Kostenki-14 and Sungir. The paternal haplogroup C-V20 can still be found in men living in modern Spain, attesting to this lineage’s longstanding presence in Western Europe. Their mitochondrial chromosomes belonged primarily to haplogroup U5. A 2023 study proposed that the Villabruna cluster emerged from the mixing in roughly equal proportions of a divergent West Eurasian ancestry with a West Eurasian ancestry closely related to the 35,000-year-old BK1653 individual from Bacho Kiro Cave in Bulgaria, with this BK1653-related ancestry also significantly (~59%) ancestral to the Věstonice cluster characteristic of eastern Gravettian producing Cro-Magnon groups, which may reflect shared ancestry in the Balkan region.” ref
“The earliest known individuals of predominantly WHG/Villabruna ancestry in Europe are known from Italy, dating to around 17,000 years ago, though an individual from El Mirón cave in northern Spain with 43% Villabruna ancestry is known from 19,000 years ago. Early WHG/Villabruna populations are associated with the Epigravettian archaeological culture, which largely replaced populations associated with the Magdalenian culture about 14,000 years ago (the ancestry of Magdalenian-associated Goyet-Q2 cluster primarily descended from the earlier Solutrean, and western Gravettian-producing groups in France and Spain). A 2023 study found that relative to earlier Western European Cro-Magnon populations like the Gravettians, that Magdalenian-associated Goyet-Q2 cluster carried significant (~30%) Villabruna ancestry even prior to the major expansion of WHG-related groups north of the Alps.” ref
“This study also found that relative to earlier members of the Villabruna cluster from Italy, WHG-related groups which appeared north of the Alps beginning around 14,000 years ago carried around 25% ancestry from the Goyet-Q2 cluster (or alternatively 10% from the western Gravettian associated Fournol cluster). This paper proposed that WHG should be named the Oberkassel cluster, after one of the oldest WHG individuals found north of the Alps. The study suggests that Oberkassel ancestry was mostly already formed before expanding, possibly around the west side of the Alps, to Western and Central Europe and Britain, where sampled WHG individuals are genetically homogeneous. This is in contrast to the arrival of Villabruna and Oberkassel ancestry to Iberia, which seems to have involved repeated admixture events with local populations carrying high levels of Goyet-Q2 ancestry. This, and the survival of specific Y-DNA haplogroup C1 clades previously observed among early European hunter-gatherers, suggests relatively higher genetic continuity in southwest Europe during this period.” ref
“The WHG were also found to have contributed ancestry to populations on the borders of Europe such as early Anatolian farmers and Ancient Northwestern Africans, as well as other European groups such as eastern hunter-gatherers. The relationship of WHGs to the EHGs remains inconclusive. EHGs are modeled to derive varying degrees of ancestry from a WHG-related lineage, ranging from merely 25% to up to 91%, with the remainder being linked to geneflow from Paleolithic Siberians (ANE) and perhaps Caucasus hunter-gatherers. Another lineage known as the Scandinavian hunter-gatherers (SHGs) were found to be a mix of EHGs and WHGs. In the Iberian Peninsula early Holocene hunter-gathers consisted of populations with a mixture of WHG and Magdalenian Cro-Magnon (GoyetQ2) ancestry.” ref
“People of the Mesolithic Kunda culture and the Narva culture of the eastern Baltic were a mix of WHG and EHG, showing the closest affinity with WHG. Samples from the Ukrainian Mesolithic and Neolithic were found to cluster tightly together between WHG and EHG, suggesting genetic continuity in the Dnieper Rapids for a period of 4,000 years. The Ukrainian samples belonged exclusively to the maternal haplogroup U, which is found in around 80% of all European hunter-gatherer samples. People of the Pit–Comb Ware culture (CCC) of the eastern Baltic were closely related to EHG. Unlike most WHGs, the WHGs of the eastern Baltic did not receive European farmer admixture during the Neolithic. Modern populations of the eastern Baltic thus harbor a larger amount of WHG ancestry than any other population in Europe.” ref
“SHGs have been found to contain a mix of WHG components who had likely migrated into Scandinavia from the south, and EHGs who had later migrated into Scandinavia from the northeast along the Norwegian coast. This hypothesis is supported by evidence that SHGs from western and northern Scandinavia had less WHG ancestry (ca 51%) than individuals from eastern Scandinavia (ca. 62%). The WHGs who entered Scandinavia are believed to have belonged to the Ahrensburg culture. EHGs and WHGs displayed lower allele frequencies of SLC45A2 and SLC24A5, which cause depigmentation, and OCA/Herc2, which causes light eye color, than SHGs.” ref
“The DNA of eleven WHGs from the Upper Palaeolithic and Mesolithic in Western Europe, Central Europe, and the Balkans was analyzed, with regards to their Y-DNA haplogroups and mtDNA haplogroups. The analysis suggested that WHGs were once widely distributed from the Atlantic coast in the West, to Sicily in the South, to the Balkans in the Southeast, for more than six thousand years. The study also included an analysis of a large number of individuals of prehistoric Eastern Europe. Thirty-seven samples were collected from Mesolithic and Neolithic Ukraine (9500-6000 BCE). These were determined to be an intermediate between EHG and SHG, although WHG ancestry in this population increased during the Neolithic. Samples of Y-DNA extracted from these individuals belonged exclusively to R haplotypes (particularly subclades of R1b1) and I haplotypes (particularly subclades of I2). mtDNA belonged almost exclusively to U (particularly subclades of U5 and U4).” ref
“A large number of individuals from the Zvejnieki burial ground, which mostly belonged to the Kunda culture and Narva culture in the eastern Baltic, were analyzed. These individuals were mostly of WHG descent in the earlier phases, but over time EHG ancestry became predominant. The Y-DNA of this site belonged almost exclusively to haplotypes of haplogroup R1b1a1a and I2a1. The mtDNA belonged exclusively to haplogroup U (particularly subclades of U2, U4 and U5). Forty individuals from three sites of the Iron Gates Mesolithic in the Balkans were also analyzed. These individuals were estimated to be of 85% WHG and 15% EHG descent. The males at these sites carried exclusively haplogroup R1b1a and I (mostly subclades of I2a) haplotypes. mtDNA belonged mostly to U (particularly subclades of U5 and U4). People of the Balkan Neolithic were found to harbor 98% Anatolian ancestry and 2% WHG ancestry. By the Chalcolithic, people of the Cucuteni–Trypillia culture were found to harbor about 20% hunter-gatherer ancestry, which was intermediate between EHG and WHG. People of the Globular Amphora culture were found to harbor ca. 25% WHG ancestry, which is significantly higher than Middle Neolithic groups of Central Europe.” ref
Scandinavian hunter-gatherer
“In archaeogenetics, the term Scandinavian hunter-gatherer (SHG) is the name given to a distinct ancestral component that represents descent from Mesolithic hunter-gatherers of Scandinavia. Genetic studies suggest that the SHGs were a mix of western hunter-gatherers (WHGs) initially populating Scandinavia from the south during the Holocene, and eastern hunter-gatherers (EHGs), who later entered Scandinavia from the north along the Norwegian coast. During the Neolithic, they admixed further with Early European Farmers (EEFs) and Western Steppe Herders (WSHs). Genetic continuity has been detected between the SHGs and members of the Pitted Ware culture (PWC), and to a certain degree, between SHGs and modern northern Europeans. The Sámi, on the other hand, have been found to be completely unrelated to the PWC.” ref
“Scandinavian hunter-gatherers (SHG) were identified as a distinct ancestral component by Lazaridis et al. (2014). A number of remains examined at Motala, Sweden, and a separate group of remains from 5,000 year-old hunter-gatherers of the Pitted Ware culture (PWC), were identified as belonging to SHG. The study found that an SHG individual from Motala (‘Motala12’) could be successfully modelled as being of c. 81% western hunter-gatherer (WHG) ancestry, and c. 19% Ancient North Eurasian (ANE) ancestry. Haak et al. (2015) examined the remains of six SHGs buried at Motala between ca. 6000 BCE and 5700 BCE. Of the four males surveyed, three carried the paternal haplogroup I2a1 or various subclades of it, while the other carried I2c. With regard to mtDNA, four individuals carried subclades of U5a, while two carried U2e1.” ref
“The study found SHGs to constitute one of the three main hunter-gatherer populations of Europe during the Mesolithic. The two other groups were WHGs and eastern hunter-gatherers (EHG). EHGs were found to be an ANE-derived population with significant admixture from a WHG-like source. SHGs formed a distinct cluster between WHG and EHG, and the admixture model proposed by Lazaridis et al. could be successfully replaced with a model that takes EHG as the source population for the ANE-like ancestry, with an admixture ratio of ~65% (WHG): ~35% (EHG). SHGs living between 6000 BCE and 3000 BCE were found to be largely genetically homogeneous, with little admixture occurring among them during this period. EHGs were found to be more closely related to SHGs than WHGs.” ref
“Mathieson et al. (2015) subjected the six SHGs from Motala to further analysis. SHGs appeared to have persisted in Scandinavia until after 5,000 years ago. The Motala SHGs were found to be closely related to WHGs. Lazaridis et al. (2016) confirmed SHGs to be a mix of EHGs (~43%) and WHGs (~57%). WHGs were modeled as descendants of the Upper Paleolithic people (Cro-Magnon) of the Grotte du Bichon in Switzerland with minor additional EHG admixture (~7%). EHGs derived c. 75% of their ancestry from ANEs. Günther et al. (2018) examined the remains of seven SHGs. All three samples of Y-DNA extracted belonged to subclades of I2. With respects to mtDNA, four samples belonged to U5a1 haplotypes, while three samples belonged U4a2 haplotypes. All samples from western and northern Scandinavia carried U5a1 haplotypes, while all the samples from eastern Scandinavia except from one carried U4a2 haplotypes.” ref
“The authors of the study suggested that SHGs were descended from a WHG population that had entered Scandinavia from the south, and an EHG population which had entered Scandinavia from the northeast along the coast. The WHGs who entered Scandinavia are believed to have belonged to the Ahrensburg culture. These WHGs and EHGs had subsequently mixed, and the SHGs gradually developed their distinct character. The SHGs from western and northern Scandinavia had more EHG ancestry (c. 49%) than individuals from eastern Scandinavia (c. 38%). The SHGs were found to have a genetic adaptation to high latitude environments, including high frequencies of low pigmentation variants and genes designed for adaptation to the cold and physical performance. SHGs displayed a high frequency of the depigmentation alleles SLC45A2 and SLC24A5, and the OCA/Herc2, which affects eye pigmentation. These genes were much less common among WHGs and EHGs. A surprising continuity was displayed between SHGs and modern populations of Northern Europe in certain respects. Most notably, the presence of the protein TMEM131 among SHGs and modern Northern Europeans was detected. This protein may be involved in long-term adaptation to the cold.” ref
“In a genetic study published in Nature Communications in January 2018, the remains of an SHG female at Motala, Sweden, between 5750 BCE and 5650 BCE was analyzed. She was found to be carrying U5a2d and “substantial ANE ancestry.” The study found that Mesolithic hunter-gatherers of the eastern Baltic also carried high frequencies of the HERC2 allele, and increased frequencies of the SLC45A2 and SLC24A5 alleles. They, however, harbored less EHG ancestry than SHGs. Genetic continuity between the SHGs and the Pitted Ware culture of the Neolithic was detected. The results further underpinned the previous suggestion that SHGs were descended from the northward migration of WHGs and a subsequent southward migration of EHGs. A certain degree of continuity between SHGs and northern Europeans was detected.” ref
“A study published in Nature in February 2018 included an analysis of a large number of individuals of prehistoric Eastern Europe. Thirty-seven samples were collected from Mesolithic and Neolithic Ukraine (9500–6000 BCE). These were determined to be an intermediate between EHG and SHG. Samples of Y-DNA extracted from these individuals belonged exclusively to R haplotypes (particularly subclades of R1b1 and R1a)) and I haplotypes (particularly subclades of I2). mtDNA belonged almost exclusively to U (particularly subclades of U5 and U4). According to Mathieson et al. (2015), 50% of Scandinavian Hunter Gatherers from Motala carried the derived variant of EDAR-V370A. This variant is typical of modern East Asian populations, and is known to affect dental morphology and hair texture, and also chin protrusion and ear morphology, as well as other facial features.” ref
“The authors did not detect East Asian ancestry in the Scandinavian Hunter Gatherers, and speculated that this gene might not have originated in East Asia, as is commonly believed. However, more recent research incorporating ancient Northeast Asian samples has confirmed that EDAR-V370A originated in Northeast Asia, and spread to West Eurasian populations such as Motala in the Holocene period. Mathieson et al. (2015) also reported: “A second surprise is that, unlike closely related western hunter-gatherers, the Motala samples have predominantly derived pigmentation alleles at SLC45A2 and SLC24A5. The study by Günther et al. (2018) further discovered that SHGs “show a combination of eye color varying from blue to light brown and light skin pigmentation. This is strikingly different from the WHGs—who have been suggested to have the specific combination of blue eyes and dark skin and EHGs—who have been suggested to be brown-eyed and light-skinned.” ref
“Four SHGs from the study yielded diverse eye and hair pigmentation predictions: one individual (SF12) was predicted to be most likely to have had dark hair and blue eyes; a second individual (Hum2) most likely had dark hair and brown eyes; a third (SF9) was predicted to have had light hair and brown eyes; and a fourth individual (SBj) was predicted to have had light hair, with the most likely hair colour being blonde, and blue eyes. Of the SHGs from Motala, four were probably dark-haired, and two others were probably light-haired, and may have been blond. In addition, all of the six SHGs from Motala had high probabilities of being blue-eyed. Both light and dark skin pigmentation alleles are found at intermediate frequencies in the Scandinavian Hunter Gatherers sampled, but only one individual had exclusively light-skin variants of two different SNPs.” ref
“The study found that depigmentation variants of genes for light skin pigmentation (SLC24A5, SLC45A2) and blue eye pigmentation (OCA2/HERC2) are found at high frequency in SHGs relative to WHGs and EHGs, which the study suggests cannot be explained simply as a result of the admixture of WHGs and EHGs. The study argues that these allele frequencies must have continued to increase in SHGs after admixture, which was probably caused by environmental adaptation to high latitudes. On the basis of archaeological and genetic evidence, the Swedish archaeologist Oscar D. Nilsson has made forensic reconstructions of both male and female SHGs.” ref
“Light skin is a human skin color that has a low level of eumelanin pigmentation as an adaptation to environments of low UV radiation. Due to the migrations of people in recent centuries, light-skinned populations today are found all over the world. Light skin is most commonly found amongst the native populations of Europe, East Asia, West Asia, Central Asia, Siberia, and North Africa as measured through skin reflectance. People with light skin pigmentation are often referred to as “white“ although these usages can be ambiguous in some countries where they are used to refer specifically to certain ethnic groups or populations.” ref
“Humans with light skin pigmentation have skin with low amounts of eumelanin, and possess fewer melanosomes than humans with dark skin pigmentation. Light skin provides better absorption qualities of ultraviolet radiation, which helps the body to synthesize higher amounts of vitamin D for bodily processes such as calcium development. On the other hand, light-skinned people who live near the equator, where there is abundant sunlight, are at an increased risk of folate depletion. As a consequence of folate depletion, they are at a higher risk of DNA damage, birth defects, and numerous types of cancers, especially skin cancer.” ref
“Humans with darker skin who live further from the tropics may have lower vitamin D levels, which can also lead to health complications, both physical and mental, including a greater risk of developing schizophrenia. These two observations form the “vitamin D–folate hypothesis,” which attempts to explain why populations that migrated away from the tropics into areas of low UV radiation evolved to have light skin pigmentation. The distribution of light-skinned populations is highly correlated with the low ultraviolet radiation levels of the regions inhabited by them. Historically, light-skinned populations almost exclusively lived far from the equator, in high-latitude areas with low sunlight intensity.” ref
“It is generally accepted that dark skin evolved as a protection against the effect of UV radiation; eumelanin protects against both folate depletion and direct damage to DNA. This accounts for the dark skin pigmentation of Homo sapiens during their development in Africa; the major migrations out of Africa to colonize the rest of the world were also dark-skinned. It is widely supposed that light skin pigmentation developed due to the importance of maintaining vitamin D3 production in the skin. Strong selective pressure would be expected for the evolution of light skin in areas of low UV radiation. After the ancestors of West Eurasians and East Eurasians diverged more than 40,000 years ago, lighter skin tones evolved independently in a subset of each of the two populations. In West Eurasians, the A111T allele of the rs1426654 polymorphism in the pigmentation gene SLC24A5 has the largest skin-lightening effect and is widespread in Europe, South Asia, Central Asia, the Near East, and North Africa.” ref
“In a 2013 study, Canfield et al. established that SLC24A5 sits in a block of haplotypes, one of which (C11) is shared by virtually all chromosomes that bear the A111T variant. This “equivalence” between C11 and A111T indicates that all people who carry this skin-lightening allele descend from a common origin: a single carrier who lived most likely “between the Middle East and the Indian subcontinent.” Canfield et al. attempted to date the A111T mutation but only constrained the age range to before the Neolithic. However, a second study from the same year (Basu Mallick et al.) estimated the coalescent age (split date) for this allele to be between ~28,000 and ~22,000 years ago.” ref
“The second most important skin-lightening factor in West Eurasians is the depigmenting allele F374 of the rs16891982 polymorphism located in the melanin-synthesis gene SLC45A2. From its low haplotype diversity, Yuasa et al. (2006) likewise concluded that this mutation (L374F) “occurred only once in the ancestry of Caucasians.” Summarising these studies, Hanel and Carlberg (2020) decided that the alleles of the two genes SLC24A5 and SLC45A2, which are most associated with lighter skin color in modern Europeans, originated in West Asia about 22,000 to 28,000 years ago and these two mutations each arose in a single carrier. This is consistent with Jones et al. (2015), who reconstructed the relationship between Near Eastern Neolithic farmers and Caucasus Hunter-Gatherers: two populations that carried the light-skin variant of SLC24A5.” ref
“Analyzing newly sequenced ancient genomes, Jones et al. estimated the split date at ~24,000 years ago and localized the separation to somewhere south of the Caucasus. However, a coalescent analysis of this allele by Crawford et al. (2017) gave a more narrowly constrained, and earlier, split date of ~29,000 years ago (with a 95% confidence window from 28,000 to 31,000 years ago). The light-skin variants of SLC24A5 and SLC45A2 were present in Anatolia by 9,000 years ago, where they became associated with the Neolithic Revolution. From here, their carriers spread Neolithic farming across Europe. Lighter skin and blond hair also evolved in the Ancient North Eurasian population.” ref
“A further wave of lighter-skinned populations across Europe (and elsewhere) is associated with the Yamnaya culture and the Indo-European migrations bearing Ancient North Eurasian ancestry and the KITLG allele for blond hair. Furthermore, the SLC24A5 gene linked with light pigmentation in Europeans was introduced into East Africa from Europe over five thousand years ago. These alleles can now be found in the San, Ethiopians, and Tanzanian populations with Afro-Asiatic ancestry. The SLC24A5 in Ethiopia maintains a substantial frequency with Semitic and Cushitic speaking populations, compared with Omotic, Nilotic, or Niger-Congo speaking groups. It is inferred that it may have arrived into the region via migration from the Levant, which is also supported by linguistic evidence. In the San people, it was acquired from interactions with Eastern African pastoralists. Meanwhile, in the case of East Asia and the Americas, a variation of the MFSD12 gene is responsible for lighter skin color. The modern association between skin tone and latitude is thus a relatively recent development.” ref
“According to Crawford et al. (2017), most of the genetic variants associated with light and dark pigmentation appear to have originated more than 300,000 years ago. African, South Asian, and Australo-Melanesian populations also carry derived alleles for dark skin pigmentation that are not found in Europeans or East Asians. Huang et al. (2021) found the existence of “selective pressure on light pigmentation in the ancestral population of Europeans and East Asians”, prior to their divergence from each other. Skin pigmentation was also found to be affected by directional selection towards darker skin among Africans, as well as lighter skin among Eurasians.” ref
“Crawford et al. (2017) similarly found evidence for selection towards light pigmentation prior to the divergence of West Eurasians and East Asians. The A111T mutation in the SLC24A5 gene predominates in populations with Western Eurasian ancestry. The geographical distribution shows that it is nearly fixed in all of Europe and most of the Middle East, extending east to some populations in present-day Pakistan and Northern India. It shows a latitudinal decline toward the Equator, with high frequencies in North Africa (80%), and intermediate (40−60%) in Ethiopia and Somalia.” ref
“Some authors have expressed caution regarding the skin pigmentation SNP predictions in early Paleolithic groups. According to Ju et al. (2021): “Relatively dark skin pigmentation in Early Upper Paleolithic Europe would be consistent with those populations being relatively poorly adapted to high-latitude conditions as a result of having recently migrated from lower latitudes. On the other hand, although we have shown that these populations carried few of the light pigmentation alleles that are segregating in present-day Europe, they may have carried different alleles that we cannot now detect. As an extreme example, Neanderthals and the Altai Denisovan individual show genetic scores that are in a similar range to Early Upper Paleolithic individuals, but it is highly plausible that these populations, who lived at high latitudes for hundreds of thousands of years, would have adapted independently to low UV levels. For this reason, we cannot confidently make statements about the skin pigmentation of ancient populations.” ref
“In 2015, it was discovered that 13,000-year-old samples of Caucasus Hunter-Gatherers (CHG) from Georgia carried the mutation and derived alleles for very fair-skinned pigmentation similar to Early Farmers (EF). This trait was said to have a relatively long history in Eurasia and risen to high frequency during the Neolithic expansion, with its origin probably predating the Last Glacial Maximum (LGM).” ref
“In the same year, a study found that genes contributing to fair skin were nearly fixed in the Anatolian Neolithic Farmers: “The second strongest signal in our analysis is at the derived allele of rs16891982 in SLC45A2, which contributes to light skin pigmentation and is almost fixed in present-day Europeans but occurred at much lower frequency in ancient populations. In contrast, the derived allele of SLC24A5 that is the other major determinant of light skin pigmentation in modern Europe, appears fixed in the Anatolian Neolithic, suggesting that its rapid increase in frequency to around 0.9 (90%) in Early Neolithic Europe was mostly due to migration.” ref
“In 2018, a study was released showing many late Mesolithic Scandinavians from 9,500 years ago in Northern Europe had blonde hair and light skin, which was in contrast to some of their contemporaries, the darker Western Hunter Gatherers (WHG). However, a 2024 paper found that phenotypically most of their studied WHG individuals carried the dark skin and blue eyes characteristic of WHGs, but some other WHGs in France they sequenced also had pale to intermediate skin pigmentation. Another entry in 2018, showed that the Eastern Hunter Gatherers (EHG), Scandinavian Hunter Gatherers (SHG), and the Baltic foragers, all had the derived alleles for light skin pigmentation.” ref
“A study on the populations of the Chalcolithic Levant (6,000-7,000 years ago), found that an allele rs1426654 in the SLC24A5 gene which is one of the most important determinants of light pigmentation in West Eurasians, was fixed for the derived variants in all Levant Chalcolithic samples, suggesting that the light skinned phenotype may have been common in the community. The individuals also had a high incidence of genomic markers associated with blue-eye color. A paper conducted by Fregel, Rosa et al. (2018) showed that in North Africa, Late Neolithic Moroccans had the European/Caucasus derived SLC24A5 mutation and other alleles and genes that predispose individuals to lighter skin and eye colours.” ref
Iberian hunter-gatherers
“The hunter-gatherers from the Iberian Peninsula carry a mix of two older types of genetic ancestry: one that dates back to the Last Glacial Maximum and was once maximized in individuals attributed to Magdalenian culture and another one that is found everywhere in western and central Europe and had replaced the Magdalenian lineage during the Early Holocene everywhere except the Iberian Peninsula,” explains Vanessa Villalba-Mouco of the Max Planck Institute for the Science of Human History, first author of the study. The researchers hope that ongoing efforts to decipher the genetic structure of late hunter-gatherer groups across Europe will help to even better understand Europe’s past and, in particular, the assimilation of a Neolithic way of life brought about by expanding farmers from the Near East during the Holocene.” ref
Ancient DNA from individuals spanning the last 8000 years helps clarify the history and prehistory of the Iberian Peninsula
“The paper published in Science focuses on slightly later time periods, and traces the population history of Iberia over the last 8000 years by analyzing ancient DNA from a huge number of individuals. The study, led by Harvard Medical School and the Broad Institute and including Haak and Villalba-Mouco, analyzed 271 ancient Iberians from the Mesolithic, Neolithic, Copper Age, Bronze Age, Iron Age, and historical periods. The large number of individuals allowed the team to make more detailed inferences about each time period than previously possible. The researchers found that during the transition to a sedentary farming life-style, hunter-gatherers in Iberia contributed subtly to the genetic make-up of newly arriving farmers from the Near East. “We can see that there must have been local mixture as the Iberian farmers also carry this dual signature of hunter-gatherer ancestry unique to Iberia,” explains Villalba-Mouco.” ref
“Between about 2500-2000 BCE, the researchers observed the replacement of 40% of Iberia’s ancestry and nearly 100% of its Y-chromosomes by people with ancestry from the Pontic Steppe, a region in what is today Ukraine and Russia. Interestingly, the findings show that in the Iron Age, “Steppe ancestry” had spread not only into Indo-European-speaking regions of Iberia but also into non-Indo-European-speaking ones, such as the region inhabited by the Basque. The researchers’ analysis suggests that present-day Basques most closely resemble a typical Iberian Iron Age population, including the influx of “Steppe ancestry,” but that they were not affected by subsequent genetic contributions that affected the rest of Iberia. This suggests that Basque speakers were equally affected genetically as other groups by the arrival of Steppe populations, but retained their language in any case. It was only after that time that they became relatively isolated genetically from the rest of the Iberian Peninsula.” ref
“Additionally, the researchers looked at historical periods, including times when Greek and later Roman settlements existed in Iberia. The researchers found that beginning at least in the Roman period, the ancestry of the peninsula was transformed by gene flow from North Africa and the eastern Mediterranean. They found that Greek and Roman settlements tended to be quite multiethnic, with individuals from the central and eastern Mediterranean and North Africa as well as locals, and that these interactions had lasting demographic as well as cultural impacts. “Beyond the specific insights about Iberia, this study serves as a model for how a high-resolution ancient DNA transect continuing into historical periods can be used to provide a detailed description of the formation of present-day populations,” explains Haak. “We hope that future use of similar strategies will provide equally valuable insights in other regions of the world.” ref
“An international team of researchers have analyzed ancient DNA from almost 300 individuals from the Iberian Peninsula, spanning more than 12,000 years, in two studies published concurrently in Current Biology and Science. The first study looked at hunter-gatherers and early farmers living in Iberia between 13,000 and 6000 years ago. The second looked at individuals from the region during all time periods over the last 8000 years. Together, the two papers greatly increase our knowledge about the population history of this unique region.” ref
“The Iberian Peninsula has long been thought of as an outlier in the population history of Europe, due to its unique climate and position on the far western edge of the continent. During the last Ice Age, Iberia remained relatively warm, allowing plants and animals – and possibly people – who were forced to retreat from much of the rest of Europe to continue living there. Similarly, over the last 8000 years, Iberia’s geographic location, rugged terrain, position on the Mediterranean coast and proximity to North Africa made it unique in comparison to other parts of Europe in its interactions with other regions. Two new studies, published concurrently in Current Biology and Science, analyze a total of almost 300 individuals who lived from about 13,000 to 400 years ago to give unprecedented clarity on the unique population history of the Iberian Peninsula.” ref
Iberian hunter-gatherers show two ancient Paleolithic lineages
“For the paper in Current Biology, led by researchers at the Max Planck Institute for the Science of Human History, researchers analyzed 11 hunter-gatherers and Neolithic individuals from Iberia. The oldest newly analyzed individuals are approximately 12,000 years old and were recovered from Balma Guilanyà in Spain. Earlier evidence had shown that, after the end of the last Ice Age, western and central Europe were dominated by hunter-gatherers with ancestry associated with an approximately 14,000-year-old individual from Villabruna, Italy. Italy is thought to have been a potential refuge for humans during the last Ice Age, like Iberia. The Villabruna-related ancestry largely replaced earlier ancestry in western and central Europe related to 19,000-15,000-year-old individuals associated with what is known as the Magdalenian cultural complex.” ref
“Interestingly, the findings of the current study show that both lineages were present in Iberian individuals dating back as far as 19,000 years ago. “We can confirm the survival of an additional Paleolithic lineage that dates back to the Late Ice Age in Iberia,” says Wolfgang Haak of the Max Planck Institute for the Science of Human History, senior author of the study. “This confirms the role of the Iberian Peninsula as a refuge during the Last Glacial Maximum, not only for fauna and flora but also for human populations.” This suggests that, far from being replaced by Villabruna-related individuals after the last Ice Age, hunter-gatherers in Iberia in fact already had ancestry from Magdalenian- and Villabruna-related sources. The discovery suggests an early connection between two potential refugia, resulting in a genetic ancestry that survived in later Iberian hunter-gatherers.” ref
Goyet-Q2 ancestry
“One of the most notable examples occurred during the Late Glacial, between the end of the LGM (~23,400 years ago) and the start of the Holocene epoch (~11,700 years ago). This shift is reflected in the ancestries associated with the ~15,090-year-old (IntCal20) Goyet Q2 individual, Belgium, and the ~14,010-year-old (IntCal20) Villabruna individual, Italy, in post-LGM Europe. We use these individuals as shorthand for the ancestries associated with them throughout the text. ‘Goyet Q2’ ancestry, which has previously been defined by the ~18,770-year-old (IntCal20) ‘El Mirón’ individual from Spain, has been identified in individuals associated with the Magdalenian culture, dating from ~20,500 to 14,000 years ago. This Goyet Q2/El Mirón ancestry has been suggested to represent a post-LGM expansion from southwestern European glacial refugia.” ref
“The ‘Villabruna’ ancestry, also broadly known as Western hunter-gatherers or WHG, consists of individuals dated from ~14,000 to 7,000 years ago associated with Epigravettian, Azilian/Federmesser, Epipalaeolithic, and Mesolithic cultures. The Villabruna ancestry is also associated with the observation that from ~14,000 years ago, all European individuals show some level of genetic affinity to present-day Near Eastern populations. The expansion in the geographic distribution of this ancestry also correlates with a period of rapid climate warming of the Late Glacial Interstadial (considered broadly equivalent to the onset of Greenland Interstadial 1 (GI-1), ~14,650 years ago) as well as cultural transitions from the Magdalenian/Late Upper Palaeolithic to the Azilian/Federmesser-Gruppen/Final Palaeolithic and has therefore been suggested to represent the movement of people into northwestern Europe after the LGM.” ref
“Interestingly, however, individuals with a mixture of Goyet Q2 and Villabruna ancestry appear in southern Europe from at least ~18,700 years ago—with the individual from El Mirón being the earliest identified thus far. The presence of individuals with admixed Goyet Q2 and Villabruna ancestry in southern Europe from the LGM onwards raises questions related to the fragmentation of populations into isolated refugia during the last Ice Age. It appears that both cultural and gene flow continued across the continent—although the nature of these processes and the mechanisms involved remain unclear. However, the presence of individuals with un-admixed Goyet Q2 ancestry in northern Europe until ~14,000 years ago also suggests some degree of sustained isolation throughout the LGM and into the Late Glacial. There is evidence of populations living in ice-marginal environments within northern Europe at the LGM and of long-distance movement of people from east to west north of the Alps, which has also been linked to the expansion of Magdalenian cultural groups. This evidence raises suggestions of Magdalenian populations with Goyet Q2 ancestry—who appear to have been cold-adapted hunter gatherers—retreating to northern Europe, perhaps due to climatic warming and the movement of prey species such as reindeer and horse. Conversely, more southerly regions such as northern Spain and Italy, where temperate prey species such as red deer persisted throughout the LGM and Late Glacial, may have provided greater ecological opportunities for population admixture.” ref
“Britain lies at the extreme northwest corner of the post-LGM expansion. With approximately two-thirds of the landmass covered by ice at the LGM and rapid deglaciation thereafter, substantial ecological and environmental change took place in the post-LGM landscape. As such, Britain offers a unique environmental context through which Late Upper Palaeolithic populations can be considered. By ~19,000 years ago, the British–Irish Ice Sheet was undergoing widespread melt, and by ~16,000 years ago, ice was absent from virtually all of England and Wales. Reindeer were present in southwest England by ~17,000 years ago, and habitats were dominated by open steppe–tundra vegetation. However, detailed consideration of Late Upper Palaeolithic sites in the United Kingdom and a series of radiocarbon dating programmes suggest that there is no evidence for post-LGM human recolonization of southwestern Britain before ~15,500 years ago.” ref
“As such, some regions of Britain were colonized before the rapid climate warming at the start of the Late Glacial Interstadial (~14,650 years ago). Accelerator mass spectrometry (AMS) dating indicates that Britain was probably recolonized at a slightly later date than adjacent regions such as the Paris Basin and the Belgian Ardennes—thereby suggesting an expansion of people across the European continent1. Interestingly, the British Magdalenian (known locally as the Creswellian) appears to be very similar (both in terms of chronology and cultural expression/typology) to the Classic Hamburgian, found in the northern Netherlands and the lowlands of northern Germany and Poland. However, understanding the expansion of post-LGM populations into and within the British Isles is hindered by a relative paucity of preserved archaeological remains suitable for dating. As such, the exact nature of human occupation of Late Upper Palaeolithic Britain remains unclear and we have relatively little knowledge of the earliest postglacial populations in Britain.” ref
“Whilst the genetics of Mesolithic, Neolithic, and Bronze Age individuals from Britain have recently been explored, no genetic data have yet been generated for British Palaeolithic individuals, due in part to the scarcity of human skeletal material available from Late Pleistocene Britain. To date, modern human skeletal remains have been recovered from only six Upper Palaeolithic sites. Nonetheless, these rare samples are crucial for our understanding of human populations across post-LGM Europe due to Britain’s location on the most northwesterly fringe of the European continent. Mesolithic British populations have been identified genetically as WHGs (Villabruna ancestry), indicating that this genetic ancestry spread to the most northwesterly area of early Holocene Europe by at least ~10,500 years ago.” ref
“What remains unclear, however, is when this ancestry first arrived in Britain and, additionally, what the genetic ancestry of Palaeolithic populations in Britain may have been. Given the previous association of Goyet Q2 ancestry with Magdalenian cultures across Europe and the similarities between the Creswellian and the Classic Hamburgian cultures, it could be hypothesized that British Late Upper Palaeolithic populations would also fall within the Goyet Q2 genetic cluster. To address these questions and expand our knowledge of the genetic makeup of Europe after the LGM, we investigate here the genetic characteristics of Late Upper Palaeolithic Britain through ancient DNA analyses of human remains from two archaeological sites in England and Wales.” ref
Anatolian hunter-gatherers
“Anatolian hunter-gatherer (AHG) is a distinct anatomically modern human archaeogenetic lineage, first identified in a 2019 study based on the remains of a single Epipaleolithic individual found in central Anatolia, radiocarbon dated to around 13,500 BCE. A population related to this individual was the main source of the ancestry of later Anatolian Neolithic Farmers (also known as Early European Farmers), who along with Western Hunter Gatherers (WHG) and Ancient North Eurasians (via Eastern Hunter Gatherers and or Western Steppe Herders) are one of the three currently known ancestral genetic contributors to present-day Europeans. The existence of this ancient population has been inferred through the genetic analysis of the remains of a man from the site of Pınarbaşı (37 ° 29’N, 33 ° 02’E), in central Anatolia, which has been dated at 13,642-13,073 cal BCE. This population is genetically differentiated from the rest of the known Pleistocene populations.” ref
“It has been discovered that populations of the Anatolian Neolithic (Anatolian Neolithic Farmers) derive most of their ancestry from the AHG, with minor gene flow from Iranian/Caucasus and Levantine sources, suggesting that agriculture was adopted in situ by these hunter-gatherers and not spread by demic diffusion into the region. The Anatolian hunter-gatherers began farming around 8300 BCE, at places such as Çayönü. Cows, sheep and goats may have been domesticated first in southern Turkey. These farmers moved into Thrace (now European Turkey) around 7000 BCE. At the autosomal level, in the Principal component analysis (PCA) the analyzed AHG individual turns out to be close to two later Anatolian populations, the Anatolian Aceramic Farmers (AAF) dating from 8300-7800 BCE, and the Anatolian Ceramic Farmers (ACF) dating from 7000-6000 BCE. The individual analyzed belongs to Y-chromosomal haplogroup C1a2 (C-V20), which has been found in some of the early WHGs, and mitochondrial haplogroup K2b. Both paternal and maternal lineages are rare in present-day Eurasian populations.” ref
“These early Anatolian farmers later replaced the European hunter-gatherer populations in Europe to a large extent, ultimately becoming the main genetic contribution to current European populations, especially those of the Mediterranean. In addition, their position in this analysis is intermediate between Natufian farmers and Western Hunter-Gatherers (WHG). This last point is confirmed by the ADMIXTURE and qp-Adm analysis and confirms the presence of hunter-gatherers of both European and Near-Eastern origins in Central Anatolia in the late Pleistocene. Mesolithic individuals from the Balkans, known as Iron Gates Hunter-Gatherers, are the most genetically similar group to the Anatolian Hunter-Gatherer lineage. Feldman et al. suggest that this affinity is not due to a genetic flow from the AHG to the ancestors of the Villabruna cluster, but on the contrary: there was a genetic flow from the ancestors of the Villabruna cluster to the ancestors of the AHG. The AHG diverged from Caucasus hunter-gatherer around 25,000 years ago.” ref
Iranian hunter-gatherers
“Despite the localization of the southern Caucasus at the outskirt of the Fertile Crescent, the Neolithisation process started there only at the beginning of the sixth millennium with the Shomutepe-Shulaveri culture of yet unclear origins. We present here genomic data for three new individuals from Mentesh Tepe in Azerbaijan, dating back to the beginnings of the Shomutepe-Shulaveri culture. Researchers have evidence that two juveniles, buried embracing each other, were brothers. We show that the Mentesh Tepe Neolithic population is the product of a recent gene flow between the Anatolian farmer-related population and the Caucasus/Iranian population, demonstrating that population admixture was at the core of the development of agriculture in the South Caucasus. By comparing Bronze Age individuals from the South Caucasus with Neolithic individuals from the same region, including Mentesh Tepe, we evidence that gene flows between Pontic Steppe populations and Mentesh Tepe-related groups contributed to the makeup of the Late Bronze Age and modern Caucasian populations.” ref
“The researcher’s results show that the high cultural diversity during the Neolithic period of the South Caucasus deserves close genetic analysis. “In the Near East, the Neolithic way of life emerged between 9000 and 7000 BCE. Several centers of Neolithisation have been identified, such as the Levant or Southern China, from which the agropastoral way of life diffused to other regions. The mechanisms of this diffusion have attracted tremendous attention for the last few decades. In some places, the Neolithic gained ground through the acculturation of local hunter-gatherers (for instance, in Anatolia or Iran); but in most regions (Europe, South-East Asia), farmer populations spread, and assimilation processes took place with a degree of admixture.” ref
“The agricultural transition profoundly changed human societies. We sequenced and analyzed the first genome (1.39x) of an early Neolithic woman from Ganj Dareh in the Zagros Mountains of Iran, a site with early evidence for an economy based on goat herding, ca. 10,000 years ago. We show that Western Iran was inhabited by a population genetically most similar to hunter-gatherers from the Caucasus, but distinct from the Neolithic Anatolian people who later brought food production into Europe. The inhabitants of Ganj Dareh made little direct genetic contribution to modern European populations, suggesting those of the Central Zagros were somewhat isolated from other populations of the Fertile Crescent. Runs of homozygosity are of a similar length to those from Neolithic farmers and shorter than those of Caucasus and Western Hunter-Gatherers, suggesting that the inhabitants of Ganj Dareh did not undergo the large population bottleneck suffered by their northern neighbors. While some degree of cultural diffusion between Anatolia, Western Iran, and other neighboring regions is possible, the genetic dissimilarity between early Anatolian farmers and the inhabitants of Ganj Dareh supports a model in which Neolithic societies in these areas were distinct.” ref
“The mechanism of Neolithisation in the South Caucasus, a region located between the Black and Caspian Seas on the southern slope of the Greater Caucasus Mountains, remains poorly understood. Mesolithic sites are known at Damjili Cave, unit 5 (Western Azerbaijan), Kmlo-2 Rock Shelter (Western Armenia), and Kotias Klde Cave (Western Georgia). Paleogenetic analyses of human bones excavated from the Kotias Klde Cave showed a genetic continuity with earlier Upper Palaeolithic (post-Last Glacial Maximum/LGM) sites but a discontinuity with pre-LGM individuals. Their genetic ancestry shares, to a certain extent, a common origin with ancient Iranian populations and differs from that of Anatolian and Levant hunter-gatherer groups, demonstrating a high genetic differentiation at this time between geographically close populations. The first settlements attributed to the Early Neolithic period belong to an aceramic culture, evidenced in several places in Central Georgia, as at Nagutni, in Western Georgia, as at Paluri, and in Western Azerbaijan at Damjili Cave, unit 4.” ref
“However, evidence of agriculture and herding remains scarce, suggesting that these sites represent a transitional phase between the Mesolithic and the Neolithic. In this context, the Shomutepe-Shulaveri culture (SSC) is the most ancient Caucasus culture with a complete Neolithic package. Found in several clusters of settlements in the northern foothills of the Lesser Caucasus, the SSC is characterized by circular mud-brick houses, domestic animals and cereals, handmade pottery, sometimes with incised and relief decoration, and obsidian and bone industries. Variants, such as the Aratashen/Aknashen culture (Ararat Plain), and other Neolithic contemporaneous cultures like the Kültepe Culture (Nakhchivan region) are also found in the South Caucasus. The slightly later Kamiltepe culture (Mil steppe culture), which probably includes the site of Polutepe, differs by its architecture, the use of flint tools instead of obsidian, and pottery-painted patterns that are rather related to Northern Iran and the Zagros.” ref
“The origins of the SSC are still discussed. Due to the rapid transition from the aceramic stage to the SSC, population continuity during the Neolithisation process is possible. However, several cultural and biological features are nonlocal. Domesticated animals, such as cattle, pigs, or goats, originate from Eastern Anatolia and from the Zagros mountains. Similarly, the glume wheat and barley recovered in the SSC sites have been domesticated elsewhere in the Middle East, even if the ancestors of the naked wheat Aegilops tauschii are found in the Caucasus and may have been local. The material culture and architecture evidence technical transfers with neighboring regions such as Southern Anatolia, Pre-Halafian, and Halafian culture of Northern Mesopotamia and Zagros11. Taken together, these data suggest a strong cultural connection, and maybe a degree of admixture, with other groups from the Fertile Crescent. Indeed, the genome-wide data for one individual coming from the same collective grave at Mentesh Tepe as the samples we analyze here, and confirmed by that of other individuals from Polutepe in the Mughan steppe or from Aknashen and Masis Blur in Armenia already showed that southern Caucasian groups are part of a cline that connected Eastern Anatolian and Zagros populations and evidence a gene flow that began around 6500 years BCE.” ref
“One of the oldest sites of the SSC, Mentesh Tepe, is located in the Tovuz district of western Azerbaijan and has been excavated between 2007 and 2015 (Fig. 1a). Several occupations were revealed, the earliest dating back to the Neolithic SSC period. At this time, the botanical assemblage is dominated by cereals, especially barley, naked wheat, and emmer, a common association during the Neolithic Southern Caucasus. Animal remains consist largely of domesticated ones (ovicaprines, cattle, pigs, dogs), and wild animals are rare. The diet of the Neolithic individuals from Mentesh Tepe relied mainly on C3-plants, such as wheat, barley, and lentils, with some evidence of freshwater fishes; the consumption of animal proteins varies between individuals. The pottery differs from classical SSC sites by being vegetal-tempered, a characteristic shared with Kamiltepe or the first occupation of the Nakhchivan site of Kültepe. As observed in many SSC sites, houses are circular and made of mudbricks, with or without the addition of straw or other organic material. Two SSC occupation phases are represented, separated in some places by a thick layer of ashes.” ref
“In a context where Neolithic burials are rare, Mentesh Tepe is exceptional for the discovery of a collective burial containing around 30 individuals (Fig. 1b), which is associated with the end of the first phase of frequentation of the site. Archaeoanthropological analysis has shown that it was a complex funerary gesture with mostly simultaneous deposits and, in contrast, some successive deposits that permitted manipulations on not completely decomposed bodies. The number of individuals in the burial, as well as their sex and age bias, suggest a dramatic event such as an epidemic, a famine, or a sudden episode, but no trace of violence has been evidenced on the bones. There is no specific orientation or position of the corpses, but some intentional arrangements are visible.” ref
“The most striking is formed by two juveniles embracing each other (Fig. 1c). Such an arrangement is rare, but other examples have been found in Neolithic and Protohistoric times, such as in Diyarbakir (Turkey, 6100 BCE) or Valdaro (Italy, 3000 BCE). Double burials are often considered as a lover’s embrace, but arguments for this explanation are often elusive. To better understand the origin of the Shomu-Shulaveri population and the structuration of this community, we performed paleogenetic studies of some of the individuals found in the collective burial. The genetic data obtained are then compared to those of another individual from the structure already published and to contemporaneous southwestern Asian genomes.” ref
Genetic structure of the Neolithic South-Caucasus
“The researchers merged our genome-wide data with the Human Origins dataset (HO-dataset), as well as with 3529 previously unrelated published ancient genomes (Supplementary Data 2). To decipher the genetic relations between the new Mentesh Tepe individuals and other ancient populations from the Caucasus, Anatolia, the Near East, and the Middle East, we performed: (1) a PCA on the modern dataset on which the ancient genomes have been projected and (2) an unsupervised ADMIXTURE analysis with the HO and the ancient dataset (Fig. 2a, b, Supplementary Fig. 2). The PCA shows that the Mentesh individuals overlap with some other previously published Neolithic or Chalcolithic individuals from the South Caucasus, but the individual from Aknashen falls a bit closer to CHG than the main neolithic cluster (Fig. 2a), and fall intermediate between the Iran Neolithic cluster and the Neolithic Anatolian Farmer group. The ADMIXTURE analysis suggests that the Mentesh individuals carry three main components: i.e., ca. 30% Neolithic Iran (Iran_N; green), 15% Levant Neolithic (PPN; pale rose) and 55% blue and pink components shared with Anatolian or European Neolithic populations (Fig. 2b).” ref
“Both analyses show that the new Mentesh Tepe individuals present a similar profile as that already published from individual MTT001 (despite different sequencing strategy) and from other Neolithic (Polutepe, Azerbaijan, Masis Blur and Aknashen, Armenia) or Chalcolithic (Alkhantepe, Azerbaijan) sites in the South Caucasus without true significant variations in percentage (Kruskal–Wallis test, p-value = 0.25), even though the Aknashen individual has the highest IranN/CHG percentage. They are also very similar to the Chalcolithic and Bronze Age Anatolian populations from Arslantepe and with other Chalcolithic and Bronze Age Anatolian populations but are quite distant from the Late Neolithic Anatolian Tell Kurdu individuals (five individuals identified as Tell_Kurdu_EC by the original publication, whom we refer as TellKurdu_LN) who instead clusters with the Neolithic Anatolian populations. Due to the lack of genomic data from South-eastern Mesopotamia, only the ancestry of groups that lived in North-eastern Mesopotamia during the PPN period and the Late Bronze Age could be considered. We observe that the South Caucasus population displays a different profile from the PPN Mesopotamian one, as it has more Anatolian affinity, but that it presents a profile similar to the LBA individual from North-eastern Mesopotamia (Nemrik9_LBA).” ref
“The agricultural transition started in a region comprising the Ancient Near East and Anatolia ~12,000 years ago with the first Pre-Pottery Neolithic villages and the first domestication of cereals and legumes. Archaeological evidence suggests a complex scenario of multiple domestications in a number of areas, coupled with examples of trade. Ancient DNA (aDNA) has revealed that this cultural package was later brought into Europe by dispersing farmers from Anatolia (so-called ‘demic’ diffusion, as opposed to non-demic cultural diffusion) ~8,400 years ago. However, a lack of aDNA from early Neolithic individuals from the Near East leaves a key question unanswered: was the agricultural transition developed by one major population group spanning the Near East, including Anatolia and the Central Zagros Mountains; or was the region inhabited by genetically diverse populations, as is suggested by the heterogeneous mode and timing of the appearance of early domesticates at different localities?” ref
“To answer this question, we sequenced the genome of an early Neolithic female from Ganj Dareh, GD13a, from the Central Zagros (Western Iran), dated to 10000-9700 years ago, a region located at the eastern edge of the Near East. Ganj Dareh is well known for providing the earliest evidence of herd management of goats beginning at 9,900 years ago. It is a classic mound site at an altitude of ~1400 m in the Gamas-Ab Valley of the High Zagros zone in Kermanshah Province, Western Iran. It was discovered in the 1960s during survey work and excavated over four seasons between 1967 and 1974. The mound, ~40 m in diameter, shows 7 to 8 m of early Neolithic cultural deposits. Five major levels were found, labeled A through E from top to bottom. Extended evidence showed a warren of rooms with evidence of under-floor inhumations within what may be burial chambers and/or disused houses. The current Minimum Number of Individuals is 116, with 56 cataloged as skeletons that had four or more bones recovered.” ref
“The individual analyzed here was part of burial 13, which contained three individuals, and was recovered in level C in 1971 from the floor of a brick-walled structure. The individual sampled, 13A (referred to as GD13a throughout the text), was a 30–50-year-old female; the other individuals in the burial unit were a second adult (13B) and an adolescent (13). The site is directly dated to 9650–9950 years ago, and it has been in intense occupation for over two to three centuries. The economy of the population was that of pastoralists with an emphasis on goat herding. Archaeobotanical evidence is limited12, but the evidence present is for two-row barley with no evidence for wheat, rye, or other domesticates. This implies that the overall economy was at a much earlier stage in the development of cereal agriculture than that found in the Levant, Anatolia, and Northern Mesopotamian basins.” ref
“The petrous bone of GD13a yielded sequencing libraries comprising 18.57% alignable human reads that were used to generate 1.39-fold genome coverage. The sequence data showed read lengths and nucleotide misincorporation patterns indicative of post-mortem damage, and had low contamination estimates (<1%, see Supplementary Fig. S3). The mitochondrion of GD13a (91.74X) was assigned to haplogroup X, most likely to the subhaplogroup X2, which has been associated with an early expansion from the Near East and has been found in early Neolithic samples from Anatolia, Hungary, and Germany.” ref
“We compared GD13a with a number of other ancient genomes and modern populations, using principal component analysis (PCA), ADMIXTURE, and outgroup f3 statistics. GD13a did not cluster with any other early Neolithic individual from Eurasia in any of the analyses. ADMIXTURE and outgroup f3 statistics identified Caucasus Hunter-Gatherers of Western Georgia, just north of the Zagros mountains, as the group genetically most similar to GD13a (Fig. 1B,C), whilst PCA also revealed some affinity with modern Central South Asian populations such as Balochi, Makrani and Brahui (Fig. 1A and Fig. S4). Also genetically close to GD13a were ancient samples from Steppe populations (Yamanya & Afanasievo) that were part of one or more Bronze age migrations into Europe, as well as early Bronze age cultures in that continent (Corded Ware), in line with previous relationships observed for the Caucasus Hunter-Gatherers.” ref
“The researchers further investigated the relationship between GD13a and Caucasus Hunter-Gatherers using D-statistics to test whether they formed a clade to the exclusion of other ancient and modern samples (Table S4). A large number of Western Eurasian samples (both modern and ancient) showed significant excess genetic affinity to the Caucasus Hunter-Gatherers, whilst none did with GD13a. Overall, these results point to GD13a having little direct genetic input into later European populations compared to its northern neighbors.” ref
“To better understand the history of the population to which GD13a belonged, we investigated the distribution of lengths of runs of homozygosity (ROH) (Fig. 2A). A bias towards a high frequency of both long and short ROH is indicative of past strong bottlenecks followed by population re-expansion. GD13a has a distribution with few long ROH (>2 Mb), similar to that of the descendants of Anatolian early farmers (represented by the European farmers NE1 and Stuttgart). In contrast, both Western and Caucasus Hunter-Gatherers have relatively more long as well as short ROH. Thus, GD13a is the descendant of a group that had relatively stable demography and did not suffer the bottlenecks that affected more northern populations.” ref
“The phenotypic attributes of GD13a are similar to the neighbouring Anatolian early farmers and Caucasus Hunter-Gatherers. Based on diagnostic SNPs, she had dark, black hair and brown eyes (see Supplementary). She lacked the derived variant (rs16891982) of the SLC45A2 gene associated with light skin pigmentation but likely had at least one copy of the derived SLC24A5 allele (rs1426654) associated with the same trait. The derived SLC24A5 variant has been found in both Neolithic farmer and Caucasus Hunter-Gatherer groups suggesting that it was already at appreciable frequency before these populations diverged. Finally, she did not have the most common European variant of the LCT gene (rs4988235) associated with the ability to digest raw milk, consistent with the later emergence of this adaptation.” ref
“It is possible that farmers related to GD13a contributed to the eastern diffusion of agriculture from the Near East that reached Turkmenistan by the 6th millennium years ago, and continued further east to the Indus Valley. However, detecting such a contribution is complicated by a later influx from Steppe populations with Caucasus Hunter-Gatherer ancestry during the Bronze Age. We tested whether the Western Eurasian component found in Indian populations can be better attributed to either of these two sources, GD13a and Kotias (a Caucasus Hunter Gatherer), using D-statistics to detect gene flow into an ancestral Indian component (represented by the Onge). Overall, for all tests where a difference could be detected, Kotias and GD13a were equally likely sources (Fig. S9 and Table S6). Whilst the attribution of part of the Western Eurasia component seen in India to Bronze Age migrations is supported by dating of last contact based on patterns of Linkage Disequilibrium, our analysis highlights the possibility that part of that component might derive from earlier contact during the eastern diffusion of agriculture.” ref
“GD13a had little direct genetic input into later European populations compared to the Caucasus Hunter-Gatherers (its northern neighbors), as demonstrated using D-statistics. This lack of connectivity with neighbouring regions might have arisen early on, since we also find that hunter-gatherers from the Caucasus show higher affinity to Western Hunter-Gatherers and early Anatolian farmers; this result suggests the possibility of gene flow between the former and these two latter groups to the exclusion of GD13a. An alternative, but not mutually exclusive, explanation for this pattern is that GD13a might have received genetic input from a source equally distant from all other European populations, and thus basal to them. The Last Glacial Maximum (LGM) made entire regions in northern Eurasia uninhabitable, and therefore a number of hunter-gatherer populations likely moved to the south. For Europe there may be a separation between Western and Eastern populations with minimal occupation of the Central European plains.” ref
“For Eastern Europe, Central Asia, and the northern Near East, glaciation itself was less a factor. In these areas, our understanding of how populations weathered the LGM is still vague at best. It has previously been suggested that differences in the frequency of long and short runs of homozygosity in ancient samples may be associated with different demographic experiences during the LGM. Neolithic farmers, with their lower frequency of short ROH, have been argued to have been relatively little affected by the LGM compared to Western and Caucasus Hunter-Gatherers which are characterised by more long ROH (>2 Mb). GD13a has a profile similar to that of the descendants of Anatolian farmers (i.e. early European farmers), suggesting that her ancestors also faced more benign conditions compared to populations further north. Superimposing the sampling locations of these groups onto climatic reconstructions from the LGM (Fig. 2B), however, does not reveal clear climatic differences among the regions. It is possible that the ancestors of the Anatolian and Ganj Dareh farmers spent the LGM in areas further south or east, which experienced milder climate.” ref
“But it is also possible that they exploited local pockets of favorable climate (refugia). Whilst high-elevation sites in the Zagros were abandoned during the LGM, there are a number of sites in the valleys that were occupied during that period and might have experienced more favorable conditions. The archaeological record indicates an eastward Neolithic expansion from the eastern regions of the Near East into Central and South Asia. Our analysis shows that both the Caucasus Hunter Gatherer Kotias and GD13a are plausible sources for the Eurasian Ancestry found in that part of Asia. Even though part of the Western Eurasian component found in India can be linked to Bronze Age migrations by dating the last contact using Linkage Disequilibrium (thus coming from the Kotias lineage), our results highlight the possibility of an older contribution from a source genetically close to GD13a (which would be hard to disentangle from the later gene flow from the Steppe).” ref
“Eventually, ancient DNA from the Indus Valley will be needed to detect conclusively whether any genetic traces were left by the eastward Neolithic expansion from the Near East, or whether this process was mostly cultural. The presence of two distinct lineages (Anatolian-like agriculturalists and Zagros mountain herders) in the Near East at the beginning of the Neolithic transition raises an interesting question regarding the independence of innovations arising at different locations. Even within the Central Zagros, economies vary greatly in their rate and pathway towards Neolithisation. Ganj Dareh, in the high Zagros, has the earliest known evidence for goat domestication, and the foothills of the Zagros mountains have also been argued to have been the site of early farming. In addition, early sites such as Sheikh-e Abad (11.650-9,600 years ago) provide evidence of early stages of barley cultivation. Were these innovations independent of similar achievements that made up the Neolithic package that North West Anatolians brought into Europe? Or were they exchanged culturally? If the latter, it would imply a cultural diffusion in the absence of much genetic interchange.” ref
“Populations of the Anatolian Neolithic derived most of their ancestry from the Anatolian hunter-gatherers (AHG), with a minor geneflow from Iranian/Caucasus and Levantine related sources, suggesting that agriculture was adopted in situ by these hunter-gatherers and not spread by demic diffusion into the region. Ancestors of AHGs and EEFs are believed to have split off from Western Hunter-Gatherers (WHGs) between 45,000 to 26,000 during the Last Glacial Maximum, and to have split from Caucasian Hunter-Gatherers (CHGs) between 25,000 to 14,000.” ref
“Genetic studies demonstrate that the introduction of farming to Europe in the 7th millennium BCE was associated with a mass migration of people from Northwest Anatolia to Southeast Europe, which resulted in the replacement of almost all (c. 98%) of the local Balkan hunter-gatherer gene pool with ancestry from Anatolian farmers. In the Balkans, the EEFs appear to have divided into two wings, who expanded further west into Europe along the Danube (Linear Pottery culture) or the western Mediterranean (Cardial Ware). Large parts of Northern Europe and Eastern Europe nevertheless remained unsettled by EEFs. During the Middle Neolithic there was a largely male-driven resurgence of WHG ancestry among many EEF-derived communities, leading to increasing frequencies of the hunter-gatherer paternal haplogroups among them.” ref
“Around 7,500 years ago. Iberian EEFs migrated into Northwest Africa, bringing farming to the region. The most common paternal haplogroup among EEFs was haplogroup G2a, while haplogroups E1b1 and R1b have also been found. Their maternal haplogroups consisted mainly of West Eurasian lineages including haplogroups H2, I, and T2, however significant numbers of central European farmers belonged to East Asian maternal lineage N9a, which is almost non-existent in modern Europeans, but common in East Asia. During the Chalcolithic and early Bronze Age, the EEF-derived cultures of Europe were overwhelmed by successive migrations of Western Steppe Herders (WSHs) from the Pontic–Caspian steppe, who carried roughly equal amounts of Eastern Hunter-Gatherer (EHG) and Caucasus Hunter-Gatherer (CHG) ancestries.” ref
“These migrations led to EEF paternal DNA lineages in Europe being almost entirely replaced with WSH-derived paternal DNA (mainly subclades of EHG-derived R1b and R1a). EEF maternal DNA (mainly haplogroup N) was also substantially replaced, being supplanted by steppe lineages, suggesting the migrations involved both males and females from the steppe. A 2017 study found that Bronze Age European with steppe ancestry had elevated EEF ancestry on the X chromosome, suggesting a sex bias, in which Steppe ancestry was inherited by more male than female ancestors. However, this study’s results could not be replicated in a follow-up study by Iosif Lazaridis and David Reich, suggesting that the authors had mis-measured the admixture proportions of their sample.” ref
“EEF ancestry remains widespread throughout Europe, ranging from about 60% near the Mediterranean Sea (with a peak of 65% in the island of Sardinia) and diminishing northwards to about 10% in northern Scandinavia. According to more recent studies the highest EEF ancestry found in modern Europeans ranges from 67% to over 80% in modern Sardinians, Italians, and Iberians, with the lowest EEF ancestry found in modern Europeans ranging around 35-40% in modern Finns, Lithuanians and Latvians. EEF ancestry is also prominent in living Northwest Africans like Moroccans and Algerians.” ref
Early European Farmers
“Early European Farmers (EEF) were a group of the Anatolian Neolithic Farmers (ANF) who brought agriculture to Europe and Northwest Africa. The Anatolian Neolithic Farmers were an ancestral component, first identified in farmers from Anatolia (also known as Asia Minor) in the Neolithic, and outside in Europe and Northwest Africa, they also existed in Iranian Plateau, South Caucasus, Mesopotamia, and Levant. Although the spread of agriculture from the Middle East to Europe has long been recognized through archaeology, it is only recent advances in archaeogenetics that have confirmed that this spread was strongly correlated with a migration of these farmers, and was not just a cultural exchange.” ref
“The earliest farmers in Anatolia derived most (80–90%) of their ancestry from the region’s local hunter-gatherers, with minor Levantine and Caucasus-related ancestry. The Early European Farmers moved into Europe from Anatolia through Southeast Europe from around 7,000 BCE, gradually spread north and westwards, and reached Northwest Africa via the Iberian Peninsula. Genetic studies have confirmed that the later Farmers of Europe generally have also a minor contribution from Western Hunter-Gatherers (WHGs), with significant regional variation. European farmer and hunter-gatherer populations coexisted and traded in some locales, although evidence suggests that the relationship was not always peaceful. Over the course of the next 4,000 years or so, Europe was transformed into agricultural communities, with WHGs being effectively replaced across Europe.” ref
“During the Chalcolithic and early Bronze Age, people who had Western Steppe Herder (WSH) ancestry moved into Europe and mingled with the EEF population; these WSH, originating from the Yamnaya culture of the Pontic steppe of Eastern Europe, probably spoke Indo-European languages. EEF ancestry is common in modern European and Northwest African populations, with EEF ancestry highest in Southern Europeans, especially Sardinians and Basque people. A distinct group of the Anatolian Neolithic Farmers spread into the east of Anatolia, and left a considerable genetic legacy in Iranian Plateau, South Caucasus, Levant (during the Pre-Pottery Neolithic B), and Mesopotamia. They also have a minor role in the ethnogenesis of WSHs of Yamnaya culture. The ANF ancestry is found in substantial levels in contemporary European, West Asian, and North African populations, and also found in Central and South Asian populations (through Bactria–Margiana Archaeological Complex and Corded Ware Culture) with more lower levels.” ref
“European hunter-gatherers were much taller than EEFs, and the replacement of European hunter-gatherers by EEFs resulted in a dramatic decrease in genetic height throughout Europe. During the later phases of the Neolithic, height increased among European farmers, probably due to increasing admixture with hunter-gatherers. During the Late Neolithic and Bronze Age, further reductions of EEF ancestry in Europe due to migrations of peoples with steppe-related ancestry is associated with further increases in height. High frequencies of EEF ancestry in Southern Europe might partly explain the shortness of Southern Europeans as compared to Northern Europeans, who carry increased levels of steppe-related ancestry. The Early European Farmers are believed to have been mostly dark-haired, dark-eyed, and light-skinned, although darker than most modern Europeans.” ref
“A study on different EEF remains throughout Europe concluded that they had “intermediate to light skin complexion.” A 2024 study found that risk alleles for mood-related phenotypes are enriched in the ancestry of Neolithic farmers. EEFs and their Anatolian forebears kept taurine cattle, pigs, sheep, and goats, as livestock, and planted cereal crops like wheat. Genetic analysis of individuals found in Neolithic tombs suggests that least some EEF peoples were patrilineal (tracing descent through the male line), with the tombs occupants mostly consisting of the male descendants of a single male common ancestor and their children, as well as their wives which were genetically unrelated to their husbands, suggesting female exogamy.” ref

To me, Animism starts in Southern Africa, then to West Europe, and becomes Totemism. Another split goes near the Russia and Siberia border becoming Shamanism, which heads into Central Europe meeting up with Totemism, which also had moved there, mixing the two which then heads to Lake Baikal in Siberia. From there this Shamanism-Totemism heads to Turkey where it becomes Paganism.

32,000-21,000 years ago, Yana Culture, at the Yana Woolly Rhinoceros Horn Site in Siberia, with genetic proximity to Ancient North Eurasian populations (Mal’ta, Afontova Gora), but also Ust-Ishim, Sunghir, and to a lesser extent Tianyuan, as well as similarities with the Clovis culture
")
ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref
“The genetic prehistory of humans in Asia, based on research using sequence data from humans who lived in Asia as early as 45,000 years ago. Genetic studies comparing present-day Australasians and Asians show that they likely derived from a single dispersal out of Africa, rapidly differentiating into three main lineages: one that persists partially in South Asia, one that is primarily found today in Australasia, and one that is widely represented across Siberia, East Asia, and Southeast Asia. Studies of ancient DNA from human remains in Asia dating from as far back as 45,000 years have greatly increased our understanding of the population dynamics leading to the current Asian populations.” ref
Ust’-Ishim man: Y-DNA haplogroupK2 and mt-DNA haplogroupR*
Tianyuan man: Y-DNA haplogroup K2b and mt-DNA haplogroup B
Yana Rhinoceros Horn Site: Y-DNA haplogroup P1 and mt-DNA haplogroup U
Sungir/Gravettian burials: Y-DNA haplogroup C1 and mt-DNA haplogroups U8c & U2
Ancient North Eurasians: Y-chromosome haplogroups P and its subclades R and Q and mt-DNA haplogroups U and R
Mal’ta–Buret’ culture: basalY-DNA haplogroup R* and mt-DNA haplogroup U
“MA-1 is the only known example of basal Y-DNA R* (R-M207*) – that is, the only member of haplogroup R* that did not belong to haplogroups R1, R2 or secondary subclades of these. The mitochondrial DNA of MA-1 belonged to an unresolved subclade of haplogroup U.” ref
“ANE ancestry has spread throughout Eurasia and the Americas in various migrations since the Upper Paleolithic, and more than half of the world’s population today derives between 5 and 42% of their genomes from the Ancient North Eurasians. Significant ANE ancestry can be found in Native Americans, as well as in Europe, South Asia, Central Asia, and Siberia. It has been suggested that their mythology may have featured narratives shared by both Indo-European and some Native American cultures, such as the existence of a metaphysical world tree and a fable in which a dog guards the path to the afterlife.” ref
Afontova Gora culture: Y-DNA haplogroup R and mt-DNA haplogroup R1b
“The bodies of two individuals, known as Afontova Gora 2 (AG2) and Afontova Gora 3 (AG3) were discovered within the complex (the name Afontova Gora 1 refers to the remains of a candid).” ref
Afontova Gora 2 “human remains”
“The human fossil remains of Afontova Gora 2 were discovered in the 1920s at Afontova Gora II and stored at the Hermitage Museum. The remains are dated to around 17,000 years ago (16,930-16,490 years ago). In 2009, researchers visited the Hermitage Museum and extracted DNA from the humerus of Afontova Gora 2. Despite significant contamination, researchers succeeded in extracting low coverage genomes. DNA analysis confirmed that the individual was male. The individual showed close genetic affinities to Mal’ta 1 (Mal’ta boy). Afontova Gora 2 also showed greater genetic affinity for the Karitiana people than for the Han Chinese. Around 1.9-2.7% of the genome was Neanderthal in origin.” ref
Afontova Gora 3 “human remains”
“In 2014, more human fossil remains were discovered at Afontova Gora II during salvage excavation before the construction of a new bridge over the Yenesei River. The remains belonged to two different females: the atlas of an adult female and the mandible and five lower teeth of a teenage girl (Afontova Gora 3), estimated to be around 14–15 years old. Initially, the new findings were presumed to be roughly contemporaneous with Afontova Gora 2. In 2017, direct AMS dating revealed that Afontova Gora 3 is dated to around 16,090 cal BCE. The mandible of Afontova Gora 3 was described as being gracile. Researchers analyzing the dental morphology of Afontova Gora 3 concluded that the teeth showed distinct characteristics with most similarities to another fossil (the Listvenka child) from the Altai-Sayan region and were neither western nor eastern. Afontova Gora 3 and Listvenka showed distinct dental characteristics that were also different from other Siberian fossils, including those from Mal’ta.” ref
“DNA was extracted from one of the teeth of Afontova Gora 3 and analyzed. Compared to Afontova Gora 2, researchers were able to obtain higher coverage genomes from Afontova Gora 3. DNA analysis confirmed that the individual was female. mtDNA analysis revealed that Afontova Gora 3 belonged to the mitochondrial Haplogroup R1b. Around 2.9-3.7% of the genome was Neanderthal in origin. In a 2016 study, researchers determined that Afontova Gora 2, Afontova Gora 3, and Mal’ta 1 (Mal’ta boy) shared common descent and were clustered together in a Mal’ta cluster. Genetically, Afontova Gora 3 is not closer to Afontova Gora 2 when compared to Mal’ta 1. When compared to Mal’ta 1, the Afontova Gora 3 lineage apparently contributed more to modern humans and is genetically closer to Native Americans.” ref
Afontova Gora 3 with Blond hair?
“Phenotypic analysis shows that Afontova Gora 3 carries the derived rs12821256 allele associated with, and likely causal for, blond hair color, making Afontova Gora 3 the earliest individual known to carry this derived allele. The allele was found in three later members of the largely ANE-derived Eastern Hunter-Gatherers populations from Samara, Motala and Ukraine c. 10,000 years ago, suggesting that it originated in the Ancient North Eurasian population before spreading to western Eurasia. The hundreds of millions of copies of this mutated alelle (a single-nucleotide polymorphism) are at the root of the classic European blond hair mutation, as massive population migrations from the Eurasian steppe, by a people who had substantial Ancient North Eurasian ancestry, entered continental Europe.” ref
The genetic proximity of Afontova Gora 3 with the Tarim mummies?
“A 2021 genetic study on the Tarim mummies found that they were primarily descended from a population represented by the Afontova Gora 3 specimen (AG3), genetically displaying “high affinity” with it. The genetic profile of the Afontova Gora 3 individual represented about 72% of the ancestry of the Tarim mummies, while the remaining 28% of their ancestry was derived from Baikal EBA (Early Bronze Age Baikal populations). The Tarim mummies are thus one of the rare Holocene populations who derive most of their ancestry from the Ancient North Eurasians (ANE, specifically the Mal’ta and Afontova Gora populations), despite their distance in time (around 14,000 years). More than any other ancient populations, they can be considered as “the best representatives” of the Ancient North Eurasians.” ref

ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref
Here are my thoughts/speculations on where I believe is the possible origin of shamanism, which may have begun sometime around 35,000 to 30,000 years ago seen in the emergence of the Gravettian culture, just to outline his thinking, on what thousands of years later led to evolved Asian shamanism, in general, and thus WU shamanism as well. In both Europe-related “shamanism-possible burials” and in Gravettian mitochondrial DNA is a seeming connection to Haplogroup U. And the first believed Shaman proposed burial belonged to Eastern Gravettians/Pavlovian culture at Dolní Věstonice in southern Moravia in the Czech Republic, which is the oldest permanent human settlement that has ever been found. It is at Dolní Věstonice where approximately 27,000-25,000 years ago a seeming female shaman was buried and also there was an ivory totem portrait figure, seemingly of her.
And my thoughts on how cultural/ritual aspects were influenced in the area of Göbekli Tepe. I think it relates to a few different cultures starting in the area before the Neolithic. Two different groups of Siberians first from northwest Siberia with U6 haplogroup 40,000 to 30,000 or so. Then R Haplogroup (mainly haplogroup R1b but also some possible R1a both related to the Ancient North Eurasians). This second group added its “R1b” DNA of around 50% to the two cultures Natufian and Trialetian. To me, it is likely both of these cultures helped create Göbekli Tepe. Then I think the female art or graffiti seen at Göbekli Tepe to me possibly relates to the Epigravettians that made it into Turkey and have similar art in North Italy. I speculate that possibly the Totem pole figurines seen first at Kostenki, next went to Mal’ta in Siberia as seen in their figurines that also seem “Totem-pole-like”, and then with the migrations of R1a it may have inspired the Shigir idol in Russia and the migrations of R1b may have inspired Göbekli Tepe.
Göbekli Tepe Shamanism
From the journal Praehistorische Zeitschrift
“Abstract: The term shamanism is widely used in archaeology to describe early belief systems. Sometimes, this has taken the form of a one-size-fits-all-explanation, without a discussion of the concept or the cultural contexts it was applied to. Recently, the Early Neolithic (9600–7000 BCE) of southwestern Asia has become a focal point of this discussion. Sites like Nevalı Çori, Göbekli Tepe, Jerf el Ahmar, Körtik Tepe, Tell Abr’3, Tell Qaramel, Wadi Faynan 16, Karahantepe and Sayburç have produced rich evidence, mostly of an iconographical nature, that seems to offer direct insights into early belief systems. The current contribution uses one of the best-researched sites, Göbekli Tepe, as a case study to develop criteria for the identification of shamanism in the archaeological record.” ref



“R1b is the most common haplogroup in Western Europe, reaching over 80% of the population in Ireland, the Scottish Highlands, western Wales, the Atlantic fringe of France, the Basque country, and Catalonia. It is also common in Anatolia and around the Caucasus, in parts of Russia, and in Central and South Asia. Besides the Atlantic and North Sea coast of Europe, hotspots include the Po valley in north-central Italy (over 70%), Armenia (35%), the Bashkirs of the Urals region of Russia (50%), Turkmenistan (over 35%), the Hazara people of Afghanistan (35%), the Uyghurs of North-West China (20%) and the Newars of Nepal (11%). R1b-V88, a subclade specific to sub-Saharan Africa, is found in 60 to 95% of men in northern Cameroon.” ref
R1b Origins & History
R1b and Paleolithic mammoth hunters
“Haplogroup R* originated in North Asia just before the Last Glacial Maximum (26,500-19,000 years ago). This haplogroup has been identified in the remains of a 24,000 year-old boy from the Altai region, in south-central Siberia (Raghavan et al. 2013). This individual belonged to a tribe of mammoth hunters that may have roamed across Siberia and parts of Europe during the Paleolithic. Autosomally this Paleolithic population appears to have contributed mostly to the ancestry of modern Europeans and South Asians, the two regions where haplogroup R also happens to be the most common nowadays (R1b in Western Europe, R1a in Eastern Europe, Central and South Asia, and R2 in South Asia).” ref
“The oldest forms of R1b (M343, P25, L389) are found dispersed at very low frequencies from Western Europe to India, a vast region where could have roamed the nomadic R1b hunter-gatherers during the Ice Age. The three main branches of R1b1 (R1b1a, R1b1b, R1b1c) all seem to have stemmed from the Middle East. The southern branch, R1b1c (V88), is found mostly in the Levant and Africa. The northern branch, R1b1a (P297), seems to have originated around the Caucasus, eastern Anatolia or northern Mesopotamia, then to have crossed over the Caucasus, from where they would have invaded Europe and Central Asia. R1b1b (M335) has only been found in Anatolia.” ref
R1b and Neolithic cattle herders
“It has been hypothesised that R1b people (perhaps alongside neighboring J2 tribes) were the first to domesticate cattle in northern Mesopotamia some 10,500 years ago. R1b tribes descended from mammoth hunters, and when mammoths went extinct, they started hunting other large game such as bisons and aurochs. With the increase of the human population in the Fertile Crescent from the beginning of the Neolithic (starting 12,000 years ago), selective hunting and culling of herds started replacing the indiscriminate killing of wild animals. The increased involvement of humans in the life of aurochs, wild boars, and goats led to their progressive taming. Cattle herders probably maintained a nomadic or semi-nomadic existence, while other people in the Fertile Crescent (presumably represented by haplogroups E1b1b, G, and T) settled down to cultivate the land or keep smaller domesticates.” ref
“The analysis of bovine DNA has revealed that all the taurine cattle (Bos taurus) alive today descend from a population of only 80 aurochs. The earliest evidence of cattle domestication dates from circa 8,500 BCE in the Pre-Pottery Neolithic cultures in the Taurus Mountains. The two oldest archaeological sites showing signs of cattle domestication are the villages of Çayönü Tepesi in southeastern Turkey and Dja’de el-Mughara in northern Iraq, two sites only 250 km away from each others. This is presumably the area from which R1b lineages started expanding – or in other words the “original homeland” of R1b.” ref
“The early R1b cattle herders would have split in at least three groups. One branch (M335) remained in Anatolia, but judging from its extreme rarity today wasn’t very successful, perhaps due to the heavy competition with other Neolithic populations in Anatolia, or to the scarcity of pastures in this mountainous environment. A second branch migrated south to the Levant, where it became the V88 branch. Some of them searched for new lands south in Africa, first in Egypt, then colonizing most of northern Africa, from the Mediterranean coast to the Sahel.” ref
“The third branch (P297), crossed the Caucasus into the vast Pontic-Caspian Steppe, which provided ideal grazing grounds for cattle. They split into two factions: R1b1a1 (M73), which went east along the Caspian Sea to Central Asia, and R1b1a2 (M269), which at first remained in the North Caucasus and the Pontic Steppe between the Dnieper and the Volga. It is not yet clear whether M73 actually migrated across the Caucasus and reached Central Asia via Kazakhstan, or if it went south through Iran and Turkmenistan. In any case, M73 would be a pre-Indo-European branch of R1b, just like V88 and M335.” ref
“R1b-M269 (the most common form in Europe) is closely associated with the diffusion of Indo-European languages, as attested by its presence in all regions of the world where Indo-European languages were spoken in ancient times, from the Atlantic coast of Europe to the Indian subcontinent, which comprised almost all Europe (except Finland, Sardinia, and Bosnia-Herzegovina), Anatolia, Armenia, European Russia, southern Siberia, many pockets around Central Asia (notably in Xinjiang, Turkmenistan, Tajikistan, and Afghanistan), without forgetting Iran, Pakistan, northern India, and Nepal. The history of R1b and R1a are intricately connected to each others.” ref
The Levantine & African branch of R1b (V88)
“Like its northern counterpart (R1b-M269), R1b-V88 is associated with the domestication of cattle in northern Mesopotamia. Both branches of R1b probably split soon after cattle were domesticated, approximately 10,500 years ago (8,500 BCE). R1b-V88 migrated south towards the Levant and Egypt. The migration of R1b people can be followed archeologically through the presence of domesticated cattle, which appear in central Syria around 8,000-7,500 BCE (late Mureybet period), then in the Southern Levant and Egypt around 7,000-6,500 BCE (e.g. at Nabta Playa and Bir Kiseiba). Cattle herders subsequently spread across most of northern and eastern Africa. The Sahara desert would have been more humid during the Neolithic Subpluvial period (c. 7250-3250 BCE), and would have been a vast savannah full of grass, an ideal environment for cattle herding.” ref
“Evidence of cow herding during the Neolithic has shown up at Uan Muhuggiag in central Libya around 5500 BCE, at the Capeletti Cave in northern Algeria around 4500 BCE. But the most compelling evidence that R1b people related to modern Europeans once roamed the Sahara is to be found at Tassili n’Ajjer in southern Algeria, a site famous pyroglyphs (rock art) dating from the Neolithic era. Some painting dating from around 3000 BCE depict fair-skinned and blond or auburn haired women riding on cows. The oldest known R1b-V88 sample in Europe is a 6,200 year-old farmer/herder from Catalonia tested by Haak et al. (2015). Autosomally this individual was a typical Near Eastern farmer, possessing just a little bit of Mesolithic West European admixture.” ref
“After reaching the Maghreb, R1b-V88 cattle herders could have crossed the Strait of Gibraltar to Iberia, probably accompanied by G2 farmers, J1 and T1a goat herders. These North African Neolithic farmers/herders could have been the ones who established the Almagra Pottery culture in Andalusia in the 6th millennium BCE.” ref
“Nowadays small percentages (1 to 4%) of R1b-V88 are found in the Levant, among the Lebanese, the Druze, and the Jews, and almost in every country in Africa north of the equator. Higher frequency in Egypt (5%), among Berbers from the Egypt-Libya border (23%), among the Sudanese Copts (15%), the Hausa people of Sudan (40%), the the Fulani people of the Sahel (54% in Niger and Cameroon), and Chadic tribes of northern Nigeria and northern Cameroon (especially among the Kirdi), where it is observed at a frequency ranging from 30% to 95% of men.” ref
“According to Cruciani et al. (2010) R1b-V88 would have crossed the Sahara between 9,200 and 5,600 years ago, and is most probably associated with the diffusion of Chadic languages, a branch of the Afroasiatic languages. V88 would have migrated from Egypt to Sudan, then expanded along the Sahel until northern Cameroon and Nigeria. However, R1b-V88 is not only present among Chadic speakers, but also among Senegambian speakers (Fula-Hausa) and Semitic speakers (Berbers, Arabs).” ref
“R1b-V88 is found among the native populations of Rwanda, South Africa, Namibia, Angola, Congo, Gabon, Equatorial Guinea, Ivory Coast, Guinea-Bissau. The wide distribution of V88 in all parts of Africa, its incidence among herding tribes, and the coalescence age of the haplogroup all support a Neolithic dispersal. In any case, a later migration out of Egypt would be improbable since it would have brought haplogroups that came to Egypt during the Bronze Age, such as J1, J2, R1a, or R1b-L23. The maternal lineages associated with the spread of R1b-V88 in Africa are mtDNA haplogroups J1b, U5, and V, and perhaps also U3 and some H subclades (=> see Retracing the mtDNA haplogroups of the original R1b people).” ref
The North Caucasus and the Pontic-Caspian steppe : the Indo-European link
“Modern linguists have placed the Proto-Indo-European homeland in the Pontic-Caspian Steppe, a distinct geographic and archeological region extending from the Danube estuary to the Ural mountains to the east and North Caucasus to the south. The Neolithic, Eneolithic, and early Bronze Age cultures in Pontic-Caspian steppe has been called the Kurgan culture (4200-2200 BCE) by Marija Gimbutas, due to the lasting practice of burying the deads under mounds (“kurgan”) among the succession of cultures in that region. It is now known that kurgan-type burials only date from the 4th millenium BCE and almost certainly originated south of the Caucasus. The genetic diversity of R1b being greater around eastern Anatolia, it is hard to deny that R1b evolved there before entering the steppe world.” ref
“Horses were first domesticated around 4600 BCE in the Caspian Steppe, perhaps somewhere around the Don or the lower Volga, and soon became a defining element of steppe culture. Nevertheless, it is unlikely that R1b was already present in the eastern steppes at the time, so the domestication of the horse should be attributed to the indigenous R1a people, or tribes belonging to the older R1b-P297 branch, which settled in eastern Europe during the Late Paleolithic or Mesolithic period. Samples from Mesolithic Samara (Haak 2015) and Latvia (Jones 2017) all belonged to R1b-P297. Autosomally these Mesolithic R1a and R1b individuals were nearly pure Mesolithic East European, sometimes with a bit of Siberian admixture, but lacked the additional Caucasian admixture found in the Chalcolithic Afanasevo, Yamna and Corded Ware samples.” ref
“It is not yet entirely clear when R1b-M269 crossed over from the South Caucasus to the Pontic-Caspian steppe. This might have happened with the appearance of the Dnieper-Donets culture (c. 5100-4300 BCE). This was the first truly Neolithic society in the Pontic-Caspian Steppe. Domesticated animals (cattle, sheep, and goats) were herded throughout the steppes and funeral rituals were elaborate. Sheep wool would play an important role in Indo-European society, notably in the Celtic and Germanic (R1b branches of the Indo-Europeans) clothing traditions up to this day.” ref
“However, many elements indicate a continuity in the Dnieper-Donets culture with the previous Mesolithic hunter-gatherers, and at the same time an influence from the Balkans and Carpathians, with regular imports of pottery and copper objects. It is, therefore, more likely that Dnieper-Donets marked the transition of indigenous R1a and/or I2a1b people to early agriculture, perhaps with an influx of Near Eastern farmers from ‘Old Europe’. Over 30 DNA samples from Neolithic Ukraine (5500-4800 BCE) were tested by Mathieson et al. (2017).” ref
“They belonged to Y-haplogroups I, I2a2, R1a, R1b1a (L754), and one R1b1a2 (L388). None of them belonged to R1b-M269 or R1b-L23 clades, which dominated during the Yamna period. Mitochondrial lineages were also exclusively of Mesolithic European origin (U4a, U4b, U4d, U5a1, U5a2, U5b2, as well as one J2b1 and one U2e1). None of those maternal lineages include typical Indo-European haplogroups, like H2a1, H6, H8, H15, I1a1, J1b1a, W3, W4 or W5 that would later show up in the Yamna, Corded Ware, and Unetice cultures. Indeed, autosomally genomes from Neolithic Ukraine were purely Mesolithic European (about 90% EHG and 10% WHG) and completely lacked the Caucasian (CHG) admxiture later found in Yamna and subsequent Indo-European cultures during the Bronze Age.” ref
“The first clearly Proto-Indo-European cultures were the Khvalynsk (5200-4500 BCE) and Sredny Stog (4600-3900 BCE) cultures in the Pontic-Caspian Steppe. This is when small kurgan burials begin to appear, with the distinctive posturing of the dead on the back with knees raised and oriented toward the northeast, which would be found in later steppe cultures as well. There is evidence of population blending from the variety of skull shapes.” ref
“Towards the end of the 5th millennium, an elite starts to develop with cattle, horses, and copper used as status symbols. It is at the turn of the Khvalynsk and Sredny Stog periods that R1b-M269’s main subclade, L23, is thought to have appeared, around 4,500 BCE. 99% of Indo-European R1b descends from this L23 clade. The other branch descended from M269 is PF7562, which is found mostly in the Balkans, Turkey, and Armenia today, and may represent an early Steppe migration to the Balkans dating from the Sredny Stog period.” ref
“Another migration across the Caucasus happened shortly before 3700 BCE, when the Maykop culture, the world’s first Bronze Age society, suddenly materialized in the north-west Caucasus, apparently out of nowhere. The origins of Maykop are still uncertain, but archeologists have linked it to contemporary Chalcolithic cultures in Assyria and western Iran. Archeology also shows a clear diffusion of bronze working and kurgan-type burials from the Maykop culture to the Pontic Steppe, where the Yamna culture developed soon afterwards (from 3500 BCE). Kurgan (a.k.a. tumulus) burials would become a dominant feature of ancient Indo-European societies and were widely used by the Celts, Romans, Germanic tribes, and Scythians, among others.” ref
“The Yamna period (3500-2500 BCE) is the most important one in the creation of Indo-European culture and society. Middle Eastern R1b-M269 people had been living and blending to some extent with the local R1a foragers and herders for over a millennium, perhaps even two or three. The close cultural contact and interactions between R1a and R1b people all over the Pontic-Caspian Steppe resulted in the creation of a common vernacular, a new lingua franca, which linguists have called Proto-Indo-European (PIE). It is pointless to try to assign another region of origin to the PIE language. Linguistic similarities exist between PIE and Caucasian and Hurrian languages in the Middle East on the one hand, and Uralic languages in the Volga-Ural region on the other hand, which makes the Pontic Steppe the perfect intermediary region.” ref
“During the Yamna period, cattle and sheep herders adopted wagons to transport their food and tents, which allowed them to move deeper into the steppe, giving rise to a new mobile lifestyle that would eventually lead to the great Indo-European migrations. This type of mass migration in which whole tribes moved with the help of wagons was still common in Gaul at the time of Julius Caesar, and among Germanic peoples in the late Antiquity.” ref
The Yamna horizon was not a single, unified culture. In the south, along the northern shores of the Black Sea coast until the the north-west Caucasus, was a region of open steppe, expanding eastward until the Caspian Sea, Siberia, and Mongolia (the Eurasian Steppe). The western section, between the Don and Dniester Rivers (and later the Danube), was the one most densely settled by R1b people, with only a minority of R1a people (5-10%). The eastern section, in the Volga basin until the Ural mountains, was inhabited by R1a people with a substantial minority of R1b people (whose descendants can be found among the Bashkirs, Turkmans, Uyghurs, and Hazaras, among others).” ref
“The northern part of the Yamna horizon was forest-steppe occupied by R1a people, also joined by a small minority of R1b (judging from Corded Ware samples and from modern Russians and Belarussians, whose frequency of R1b is from seven to nine times lower than R1a). The western branch would migrate to the Balkans and Greece, then to Central and Western Europe, and back to their ancestral Anatolia in successive waves (Hittites, Phrygians, Armenians, etc.). The eastern branch would migrate to Central Asia, Xinjiang, Siberia, and South Asia (Iran, Pakistan, India). The northern branch would evolve into the Corded Ware culture and disperse around the Baltic, Poland, Germany, and Scandinavia.” ref
The Maykop culture, the R1b link to the Steppe?
“The Maykop culture (3700-2500 BCE) in the north-west Caucasus was culturally speaking a sort of southern extension of the Yamna horizon. Although not generally considered part of the Pontic-Caspian steppe culture due to its geography, the North Caucasus had close links with the steppes, as attested by numerous ceramics, gold, copper, and bronze weapons and jewelry in the contemporaneous cultures of Mikhaylovka, Sredny Stog, and Kemi Oba. The link between the northern Black Sea coast and the North Caucasus is older than the Maykop period. Its predecessor, the Svobodnoe culture (4400-3700 BCE), already had links to the Suvorovo-Novodanilovka and early Sredny Stog cultures. The even older Nalchik settlement (5000-4500 BCE) in the North Caucasus displayed a similar culture as Khvalynsk in the Caspian Steppe and Volga region. This may be the period when R1b started interracting and blending with the R1a population of the steppes.” ref
“The Yamna and Maykop people both used kurgan burials, placing their deads in a supine position with raised knees and oriented in a north-east/south-west axis. Graves were sprinkled with red ochre on the floor, and sacrificed domestic animal buried alongside humans. They also had in common horses, wagons, a heavily cattle-based economy with a minority of sheep kept for their wool, use of copper/bronze battle-axes (both hammer-axes and sleeved axes), and tanged daggers. In fact, the oldest wagons and bronze artifacts are found in the North Caucasus, and appear to have spread from there to the steppes.” ref
‘Maykop was an advanced Bronze Age culture, actually one of the very first to develop metalworking, and therefore metal weapons. The world’s oldest sword was found at a late Maykop grave in Klady kurgan 31. Its style is reminiscent of the long Celtic swords, though less elaborated. Horse bones and depictions of horses already appear in early Maykop graves, suggesting that the Maykop culture might have been founded by steppe people or by people who had close link with them. However, the presence of cultural elements radically different from the steppe culture in some sites could mean that Maykop had a hybrid population.” ref
“Without DNA testing it is impossible to say if these two populations were an Anatolian R1b group and a G2a Caucasian group, or whether R1a people had settled there too. The two or three ethnicities might even have cohabited side by side in different settlements. The one typical Caucasian Y-DNA lineage that does follow the pattern of Indo-European migrations is G2a-L13, which is found throughout Europe, Central Asia, and South Asia. In the Balkans, the Danube basin, and Central Europe its frequency is somewhat proportional to the percentage of R1b.” ref
“Maykop people are the ones credited for the introduction of primitive wheeled vehicles (wagons) from Mesopotamia to the Steppe. This would revolutionise the way of life in the steppe, and would later lead to the development of (horse-drawn) war chariots around 2000 BCE. Cavalry and chariots played an vital role in the subsequent Indo-European migrations, allowing them to move quickly and defeat easily anybody they encountered. Combined with advanced bronze weapons and their sea-based culture, the western branch (R1b) of the Indo-Europeans from the Black Sea shores are excellent candidates for being the mysterious Sea Peoples, who raided the eastern shores of the Mediterranean during the second millennium BCE.” ref
“The rise of the IE-speaking Hittites in Central Anatolia happened a few centuries after the disappearance of the Maykop and Yamna cultures. Considering that most Indo-European forms of R1b found in Anatolia today belong to the R1b-Z2103 subclade, it makes little doubt that the Hittites came to Anatolia via the Balkans, after Yamna/Maykop people invaded Southeast Europe. The Maykop and Yamna cultures were succeeded by the Srubna culture (1600-1200 BCE), possibly representing an advance of R1a-Z282 people from the northern steppes towards the Black Sea shores, filling the vacuum left by the R1b tribes who migrated to Southeast Europe and Anatolia.” ref
The Siberian & Central Asian branch
“When R1b crossed the Caucasus in the Late Neolithic, it split into two main groups. The western one (L51) would settle the eastern and northern of the Black Sea. The eastern one (Z2103) migrated to the Don-Volga region, where horses were domesticated circa 4600 BCE. R1b probably mixed with indigenous R1a people and founded the Repin culture (3700-3300 BCE) a bit before the Yamna culture came into existence in the western Pontic Steppe. R1b would then have migrated with horses along the Great Eurasian Steppe until the Altai mountains in East-Central Asia, where they established the Afanasevo culture (c. 3600-2400 BCE). Afanasevo people might be the precursors of the Tocharian branch of Indo-European languages. In 2014, Clément Hollard of Strasbourg University tested three Y-DNA samples from the Afanasevo culture and all three turned out to belong to haplogroup R1b, including two to R1b-M269.” ref
“The R1b people who stayed in the Volga-Ural region were probably the initiators of the Poltavka culture (2700-2100 BCE), then became integrated into the R1a-dominant Sintashta-Petrovka culture (2100-1750 BCE) linked to the Indo-Aryan conquest of Central and South Asia (=> see R1a for more details).” ref
“Nowadays in Russia R1b is found at higher frequencies among ethnic minorities of the Volga-Ural region (Udmurts, Komi, Mordvins, Tatars) than among Slavic Russians. R1b is also present in many Central Asian populations, the highest percentages being observed among the Uyghurs (20%) of Xinjiang in north-west China, the Yaghnobi people of Tajikistan (32%), and the Bashkirs (47%, or 62.5% in the Abzelilovsky district) of Bashkortostan in Russia (border of Kazakhstan).” ref
“R1b-M73, found primarily in North Asia (Altai, Mongolia), Central Asia, and the North Caucasus is thought to have spread during the Neolithic from the Middle East to Central and North Asia, and therefore can be considered to be pre-Indo-European.” ref
The European & Middle Eastern branch
“The Indo-Europeans’s bronze weapons and the extra mobility provided by horses would have given them a tremendous advantage over the autochthonous inhabitants of Europe, namely the native haplogroup C1a2, F and I (descendants of Cro-Magnon) and the early Neolithic herders and farmers (G2a, H2, E1b1b, and T1a). This allowed R1a and R1b to replace most of the native male lineages (=> see How did R1b come to replace most of the older lineages in Western Europe?), although female lineages seem to have been less affected.” ref
“A comparison with the Indo-Iranian invasion of South Asia shows that 40% of the male linages of northern India are R1a, but less than 10% of the female lineages could be of Indo-European origin. The impact of the Indo-Europeans was more severe in Europe because European society 4,000 years ago was less developed in terms of agriculture, technology (no bronze weapons), and population density than that of the Indus Valley civilization.” ref
“This is particularly true of the native Western European cultures where farming arrived much later than in the Balkans or Central Europe. Greece, the Balkans, and the Carpathians were the most advanced of European societies at the time and were the least affected in terms of haplogroup replacement. neolithic lineages survived better in regions that were more difficult to reach or less hospitable to horse breeders, like the Alps, the Dinaric Alps, the Apennines, and Sardinia.” ref
The Conquest of “Old Europe” and Central Europe (4200-2500 BCE)
“The first forays of Steppe people into the Balkans happened between 4200 BCE and 3900 BCE, when cattle herders equipped with horse-drawn wagons crossed the Dniester and Danube and apparently destroyed the towns of the Gumelnița, Varna and Karanovo VI cultures in Eastern Romania and Bulgaria. A climatic change resulting in colder winters during this exact period probably pushed steppe herders to seek milder pastures for their stock, while failed crops would have led to famine and internal disturbance within the Danubian and Balkanic communities. The ensuing Cernavodă culture (Copper Age, 4000-3200 BCE), Coțofeni/Usatovo culture (Copper to Bronze Age, 3500-2500 BCE), Ezero culture (Bronze Age, 3300-2700 BCE), in modern Romania, seems to have had a mixed population of steppe immigrants and people from the old tell settlements. These Steppe immigrants were likely a mixture of both R1a and R1b lineages, with a probably higher percentage of R1a than later Yamna-era invasions.” ref
“The Steppe invaders would have forced many Danubian farmers to migrate to the Cucuteni-Trypillian towns in the eastern Carpathians, causing a population boom and a north-eastward expansion until the Dnieper valley, bringing Y-haplogroups G2a, I2a1 (probably the dominant lineage of the Cucuteni-Trypillian culture), E1b1b, J2a, and T1a in what is now central Ukraine. This precocious Indo-European advance westward was fairly limited, due to the absence of Bronze weapons and an organized army at the time, and was indeed only possible thanks to climatic catastrophes which reduced the defenses of the towns of Old Europe. The Carphatian, Danubian, and Balkanic cultures were too densely populated and technologically advanced to allow for a massive migration.” ref
“In comparison, the forest-steppe R1a people successfully penetrated into the heart of Europe with little hindrance, due to the absence of developed agrarian societies around Poland and the Baltic. The Corded Ware culture (3200-1800 BCE) was a natural northern and western expansion of the Yamna culture, reaching as far west as Germany and as far north as Sweden and Norway. DNA analysis from the Corded Ware confirmed the presence of R1a and R1b in Poland c. 2700 BCE and R1a central Germany around 2600 BCE. The Corded Ware tribes expanded from the northern fringe of the Yamna culture where R1a lineages were prevalent over R1b ones.” ref
“The expansion of R1b people into Old Europe was slower, but proved inevitable. In 2800 BCE, by the time the Corded Ware had already reached Scandinavia, the Bronze Age R1b cultures had barely moved into the Pannonian Steppe. They established major settlements in the Great Hungarian Plain, the most similar habitat to their ancestral Pontic Steppes. Around 2500 BCE, the western branch of Indo-European R1b were poised for their next major expansion into modern Germany and Western Europe. By that time, the R1b immigrants had blended to a great extent with the indigenous Mesolithic and Neolithic populations of the Danubian basin, where they had now lived for 1,700 years.” ref
“The strongly partriarchal Indo-European elite remained almost exclusively R1b on the paternal side, but absorbed a high proportion of non-Indo-European maternal lineages. Hybridised, the new Proto-Indo-European R1b people would have lost most of their remaining Proto-Europoid or Mongolid features inherited from their Caspian origins (which were still clearly visible in numerous individuals from the Yamna period). Their light hair, eye, and skin pigmentation, once interbred with the darker inhabitants of Old Europe, became more like that of modern Southern Europeans.” ref
“The R1a people of the Corded Ware culture would come across far less populous societies in Northern Europe, mostly descended from the lighter Mesolithic population, and therefore retained more of their original pigmentation (although facial traits evolved considerably in Scandinavia, where the I1 inhabitants were strongly dolicocephalic and long-faced, as opposed to the brachycephalic and broad-faced Steppe people).” ref
The Conquest of Western Europe (2500-1200 BCE)
“The R1b conquest of Europe happened in two phases. For nearly two millennia, starting from circa 4200 BCE, Steppe people limited their conquest to the rich Chalcolithic civilizations of the Carpathians and the Balkans. These societies possessed the world’s largest towns, notably the tell settlements of the Cucuteni-Tripolye culture. Nothing incited the R1b conquerors to move further into Western Europe at such an early stage, because most of the land north and west of the Alps was still sparsely populated woodland. The Neolithic did not reach the British Isles and Scandinavia before circa 4000 BCE. Even northern France and most of the Alpine region had been farming or herding for less than a millennium and were still quite primitive compared to Southeast Europe and the Middle East.” ref
“North-west Europe remained a tribal society of hunter-gatherers practicing only limited agriculture for centuries after the conquest of the Balkans by the Indo-Europeans. Why would our R1b “conquistadors” leave the comfort of the wealthy and populous Danubian civilizations for the harsh living conditions that lie beyond? Bronze Age people coveted tin, copper, and gold, of which the Balkans had plenty, but that no one had yet discovered in Western Europe.” ref
“R1b-L51 is thought to have arrived in Central Europe (Hungary, Austria, Bohemia) around 2500 BCE, approximately two millennia after the shift to the Neolithic lifestyle in these regions. Agrarian towns had started to develop. Gold and copper had begun to be mined. The prospects of a conquest were now far more appealing.” ref
The archeological and genetic evidence (distribution of R1b subclades) point at several consecutive waves towards eastern and central Germany between 2800 BCE and 2300 BCE. The Unetice culture was probably the first culture in which R1b-L11 lineages played a major role. It is interesting to note that the Unetice period happen to correspond to the end of the Maykop (2500 BCE) and Kemi Oba (2200 BCE) cultures on the northern shores of the Black Sea, and their replacement by cultures descended from the northern steppes.” ref
“It can therefore be envisaged that the (mostly) R1b population from the northern half of the Black Sea migrated westward due to pressure from other Indo-European people (R1a) from the north, for example that of the burgeoning Proto-Indo-Iranian branch, linked to the contemporary Poltavka and Abashevo cultures.” ref
“It is doubtful that the Bell Beaker culture (2900-1800 BCE) in Western Europe was already Indo-European because its attributes are in perfect continuity with the native Megalithic cultures. The Beaker phenomenon started during the Late Neolithic and Early Chalcolithic in Portugal and propagated to the north-east towards Germany. During the same period Bronze Age Steppe cultures spread from Germany in the opposite direction towards Iberia, France, and Britain, progressively bringing R1b lineages into the Bell Beaker territory.” ref
“It is more likely that the beakers and horses found across Western Europe during that period were the result of trade with neighboring Indo-European cultures, including the first wave of R1b into Central Europe. It is equally possible that the Beaker people were R1b merchants or explorers who traveled across Western Europe and brought back tales of riches poorly defended by Stone Age people waiting to be to be conquered. This would have prompted a full-scale Indo-European (R1b) invasion from about 2500 BCE in Germany, reaching the Atlantic (north of the Pyrenees at least) around 2200 BCE.” ref
“Ancient DNA tests conducted by Lee et al. (2012), Haak et al. (2015), and Allentoft et al. (2015) have all confirmed the presence of R1b-L51 (and deeper subclades such as P312 and U152) in Germany from the Bell Beaker period onwards, but none in earlier cultures. German Bell Beaker R1b samples only had about 50% of Yamna autosomal DNA and often possessed Neolithic non-Steppe mtDNA, which confirms that R1b invaders took local wives as they advanced westward. Another study by Olalde et al. (2017) confirmed that Iberian Bell Beakers were genetically distinct from the previously tested German samples.” ref
“None of the Spanish or Portuguese individuals associated with Bell Beaker pottery possessed any Steppe admixture, and none belonged to the Indo-European haplogroup R1b-L23 or its subclades. Instead, they belonged to typical Megalithic lineages like G2a, I2a1, I2a2, and the Neolithic R1b-V88. The paper also confirmed a high frequency of R1b-L51 lineages in central Europe during the Beall Beaker period. In Britain, Megalithic individuals belonged exclusively to Y-haplogroup I2 (mostly I2a2 and I2a1b-L161), but were entirely replaced by R1b-L51 (mostly L21 clade) in the Early Bronze Age.” ref
“This means that the Bell Beaker culture was not associated with one particular ethnic group. Beaker pottery originated in Megalithic Iberia, but then spread to France and central Europe and was used by invading R1b-L51 Steppe people, who brought it with them to the British Isles, while wiping out most of the indigenous Megalithic population. There was, therefore, no ‘Bell Beaker people’, but just various populations trading and using Beaker pots during that period.” ref
“DNA samples from the Unetice culture (2300-1600 BCE) in Germany, which emerged less than two centuries after the appearance of the first R1b-L51 individuals in the late Bell Beaker Germany, had a slightly higher percentage of Yamna ancestry (60~65%) and of Yamna-related mtDNA lineages, which indicates a migration of both Steppe men and women. That would explain why archeological artifacts from the Unetice culture are clearly Yamna-related (i.e. Indo-European), as they abruptly introduced new technologies and a radically different lifestyle, while the Bell Beaker culture was in direct continuity with previous Neolithic or Chalcolithic cultures.” ref
“R1b men may simply have conquered the Bell Beaker people and overthrown the local rulers without obliterating the old culture due to their limited numbers. Taking the analogy of the Germanic migrations in the Late Antiquity, the R1b invasion of the Bell Beaker period was more alike to that of the Goths, Burgunds and Vandals, who all migrated in small numbers, created new kingdoms within the Roman empire, but adopted Latin language and Roman culture. In contrast, the Corded Ware and Unetice culture involved large-scale migrations of Steppe people, who imposed their Indo-European language and culture and conquered people, just like the Anglo-Saxons or the Bavarians did in the 5th century.” ref
“The cultures that succeeded to Unetice in Central Europe, chronologically the Tumulus culture (1600-1200 BCE), Urnfield culture (1300-1200 BCE), and Hallstatt culture (1200-750 BCE) cultures remained typically Indo-European. The Hallstatt culture, centered around the Alps, is considered the first classical Celtic culture in Europe. It quickly expanded to France, Britain, Iberia, northern Italy, and the Danube valley, probably spreading for the first time Celtic languages, although not bronze technology nor R1b lineages, which had both already spread over much of western Europe during the Bell Beaker period. => See also Metal-mining and stockbreeding explain R1b dominance in Atlantic fringe.” ref
“The linguistic gap between pre-IE vernaculars and IE languages was about as big as between modern English and Chinese. English, Greek, Russian, and Hindi are all related IE languages and therefore easier to learn for IE speakers than non-IE languages like Chinese, Arabic, or Hungarian. From a linguistic point of view, only a wide-scale migration of IE speakers could explain the thorough adoption of IE languages in Western Europe – leaving only Basque as a remnant of the Neolithic languages.” ref
“Besides pottery, archaeology provides ample evidence that the early Bronze Age in Central and Western Europe coincides with a radical shift in food production. Agriculture experiences an abrupt reduction in exchange for an increased emphasis on domesticates. This is also a period when horses become more common and cow milk is being consumed regularly. The overall change mimics the Steppic way of life almost perfectly. Even after the introduction of agriculture around 5200 BCE, the Bug-Dniester culture and later Steppe cultures were characterized by an economy dominated by herding, with only limited farming. This pattern expands into Europe exactly at the same time as bronze working.” ref
“Religious beliefs and arts undergo a complete reversal in Bronze Age Europe. Neolithic societies in the Near East and Europe had always worshipped female figurines as a form of fertility cult. The Steppe cultures, on the contrary, did not manufacture female figurines. As bronze technology spreads from the Danube valley to Western Europe, symbols of fertility and fecundity progressively disappear and are replaced by sculptures of domesticated animals.” ref
“Another clue that Indo-European Steppe people came in great number to Central and Western Europe is to be found in burial practices. Neolithic Europeans either cremated their dead (e.g. Cucuteni-Tripolye culture) or buried them in collective graves (this was the case of Megalithic cultures). In the Steppe, each person was buried individually, and high-ranking graves were placed in a funeral chamber and topped by a circular mound. The body was typically accompanied by weapons (maces, axes, daggers), horse bones, and a dismantled wagon (or later chariot).” ref
“These characteristic burial mounds are known as kurgans in the Pontic Steppe. Men were given more sumptuous tombs than women, even among children, and differences in hierarchy are obvious between burials. The Indo-Europeans had a strongly hierarchical and patrilinear society, as opposed to the more egalitarian and matrilinear cultures of Old Europe. The proliferation of ststus-conscious male-dominant kurgans (or tumulus) in Central Europe during the Bronze Age is a clear sign that the ruling elite had now become Indo-European.” ref
“The practice also spread to central Asia and southern Siberia, two regions where R1a and R1b lineages are found nowadays, just like in Central Europe. The ceremony of burial is one of the most emotionally charged and personal aspect of a culture. It is highly doubtful that people would change their ancestral practice “just to do like the neighbours”. In fact, different funerary practices have co-existed side by side during the European Neolithic and Chalcolithic. The ascendancy of yet another constituent of the Pontic Steppe culture in the rest of Europe, and in this case one that does not change easily through contact with neighbours, adds up to the likelihood of a strong Indo-European migration.” ref
“The adoption of some elements of a foreign culture tends to happen when one civilization overawes the adjacent cultures by its superiority. This process is called ‘acculturation’. However, there is nothing that indicates that the Steppe culture was so culturally superior as to motivate a whole continent, even Atlantic cultures over 2000 km away from the Pontic Steppe, to abandon so many fundamental symbols of their own ancestral culture, and even their own language. In fact, Old Europe was far more refined in its pottery and jewelry than the rough Steppe people. The Indo-European superiority was cultural but military, thanks to horses, bronze weapons, and an ethic code valuing individual heroic feats in war (these ethic values are known from the old IE texts, like the Rig Veda, Avesta, or the Mycenaean and Hittite literature).” ref
“After linguistics and archaeology, the third category of evidence comes from genetics itself. It had first been hypothesized that R1b was native to Western Europe, because this is where it was most prevalent. It has since been proven that R1b haplotypes displayed higher microsatellite diversity in Anatolia and in the Caucasus than in Europe. European subclades are also more recent than Middle Eastern or Central Asian ones. The main European subclade, R-P312/S116, only dates back to approximately 3500 to 3000 BCE.” ref
“It does not mean that the oldest common ancestor of this lineage arrived in Western Europe during this period, but that the first person who carried the mutation R-P312/S116 lived at least 5,000 years ago, assumably somewhere in the lower Danube valley or around the Black Sea. In any case, this timeframe is far too recent for a Paleolithic origin or a Neolithic arrival of R1b. The discovery of what was thought to be “European lineages” in Central Asia, Pakistan, and India hit the final nail on the coffin of a Paleolithic origin of R1b in Western Europe, and confirmed the Indo-European link.” ref
“All the elements concur in favor of a large scale migration of Indo-European speakers (possibly riding on horses) to Western Europe between 2500 to 2100 BCE, contributing to the replacement of the Neolithic or Chalcolithic lifestyle by an inherently new Bronze Age culture, with simpler pottery, less farming, more herding, new rituals (single graves) and new values (patrilinear society, warrior heroes) that did not evolve from local predecessors.” ref







1")
Haplogroup migrations related to the Ancient North Eurasians: I added stuff to this map to help explain.
People reached Lake Baikal Siberia around 25,000 years ago. They (to Damien) were likely Animistic Shamanists who were also heavily totemistic as well. Being animistic thinkers they likely viewed amazing things in nature as a part of or related to something supernatural/spiritual (not just natural as explained by science): spirit-filled, a sprit-being relates to or with it, it is a sprit-being, it is a supernatural/spiritual creature, or it is a great spirit/tutelary deity/goddess-god. From there comes mythology and faith in things not seen but are believed to somehow relate or interact with this “real world” we know exists.
Both areas of Lake Baikal, one on the west side with Ancient North Eurasian culture and one on the east side with Ancient Northern East Asian culture (later to become: Ancient Northeast Asian culture) areas are the connected areas that (to Damien) are the origin ancestry religion area for many mythologies and religious ideas of the world by means of a few main migrations and many smaller ones leading to a distribution of religious ideas that even though are vast in distance are commonly related to and centering on Lake Baikal and its surrounding areas like the Amur region and Altai Mountains region.
To an Animistic Thinker: “Things are not just as they seem, they may have a spirit, or spirit energy relates to them”
To a Totemistic Thinker: “Things are not just as they seem, they may have a spirit, or spirit energy relates to them; they may have religio-cultural importance.”
“Ancient North Eurasian population had Haplogroups R, P, U, and Q DNA types: defined by maternal West-Eurasian ancestry components (such as mtDNA haplogroup U) and paternal East-Eurasian ancestry components (such as yDNA haplogroup P1 (R*/Q*).” ref


“Lighter skin and blond hair evolved in the Ancient North Eurasian (ANE) population. The SLC24A5 gene’s derived threonine or Ala111Thr allele (rs1426654) has been shown to be a major factor in the light skin tone of Europeans. Possibly originating as long as 19,000 years ago, it has been the subject of selection in the ancestors of Europeans as recently as within the last 5,000 years, and is fixed in modern European populations.” ref, ref
DNA-researcher: It’s not ‘woke’ to portray prehistoric Europeans with dark skin.
“It’s evolution. Ancient DNA analyses suggest that prehistoric Europeans looked different from modern Europeans today, but some people find that hard to accept. There was an artistic picture of an almost 6,000-year-old, girl who was walking along Lolland’s south coast and spits a piece of birch tar into the reeds. It didn’t taste great, but it helped to soothe her toothache. Fast forward 6,000 years, Danish archaeologists working on the Fehmarnbelt project stumble across the piece and recognize it for what it is: an almost 6,000-year-old piece of chewing gum. This ancient piece of gum is now on display at the Museum Lolland-Falster in southern Denmark among an amazing collection of Stone Age artifacts uncovered during the excavations. If you have not been, it is well worth a visit. In 2019, my research team at the University of Copenhagen managed something quite remarkable: We succeeded in extracting DNA from the gum and used it to reconstruct the girl’s entire genome — the first time anyone had sequenced an ancient human genome from anything other than skeletal remains. As the gum had been found on Lolland, we affectionately nicknamed her ‘Lola’.” ref
Stone-age girl in social media ‘shitstorm’
“The story of Lola and her chewing gum made headlines around the world when we published the genome in 2019 and then, suddenly, in the summer of 2023, Lola was back in the news, caught up in a media ‘shitstorm’. The ‘shitstorm’ first gathered pace on X, the platform formerly known as Twitter, and escalated to the point where the museum had to defend itself on national TV. Even the Danish newspaper ‘Ekstrabladet’ felt they had to comment and gave their opinion in a passionate editorial. So, what happened? These things are difficult to reconstruct, but evidently some people who had seen the image of Lola thought that she looked “way too dark” and accused us—and the museum—of ‘blackwashing’ the past. I suppose this episode says more about our own biases than anything else, and I would like to take this opportunity to explain why we portrayed Lola the way we did and what this tells us about the evolution of skin color in this part of the world.” ref
What we know about Lola
“First a disclaimer, we do not know exactly how old Lola was when she spat that chewing gum into the water. But based on her genome and other DNA trapped in the gum, we learned a lot of other things about her and her world. For example, we learned that she was a hunter-gatherer who lived off wild resources like fish, nuts, and wild game. At the time, small farming communities started to appear in other parts of Europe, but from what we can tell Lola and her kin still lived — as her ancestors had done for thousands of years before her — as hunter-gatherers. We also learned that she likely had dark skin, dark hair, and blue eyes. But how do we know that?” ref
The genetics of human skin pigmentation
“Skin color is a highly heritable and polygenic trait, meaning that it is influenced by multiple genes and their interactions with one another. One of the most well-known genes associated with skin pigmentation is the melanocortin 1 receptor gene (MC1R), but there are dozens more that have been reported to be involved in the pigmentation process. Most of these genes influence skin color by regulating the production of melanin, a dark pigment that protects from the deleterious effects of UV radiation. Basically, the more melanin you have in your skin, the darker it will be, and the more sun your skin can tolerate before you get sunburn. Eye and hair color are determined in a similar way, but the mechanisms that control the production of melanin in the eyes and hair are quite complex and independent processes. That is why it is possible to end up with different combinations of traits, such as the dark hair and blue eyes that are often seen in Europeans today, or the light hair and brown eyes that are common for Solomon Islanders, for example.” ref
How do we know what Lola looked like?
“Because the genes involved in pigmentation have been well studied, it is possible to predict the skin, eye, and hair color of an individual based on their genotype with a certain probability, something that is routinely done in forensic investigations. In practice, this works by checking which variants of a gene are present and what phenotype they are associated with. The more genes we can include in this analysis, the more confident we can be that our prediction is correct. In Lola’s case, we studied 41 gene variants across her genome that have been associated with skin, hair, and eye color in humans, and concluded that she likely had this unusual (at least for today) combination of dark skin, dark hair, and blue eyes.” ref
A common look in prehistoric Europe
“It is difficult to know exactly what people looked like 10,000 years ago. But based on ancient DNA studies, it appears that Lola’s ‘look’ was much more common in prehistoric Europe than it is today. Thanks to advances in ancient DNA sequencing, we now have the genomes of dozens of Upper Palaeolithic and Mesolithic (i.e. the period between around 50,000 and 5,000 years before present in Europe) individuals from Western Europe. And interestingly they all seem to lack the skin-lightening variants that are so common in Europeans today, indicating that they had dark skin. This is true for ‘Cheddar Man’ who lived around 10,000 years ago in southern England, as well as dozens of other Upper Palaeolithic and Mesolithic hunter-gatherer individuals from France, northern Italy, Spain, the Baltic, and other parts of Europe. Like skin color, eye color is also a fairly complex trait, involving the interaction of many different genes. Therefore, eye color is fairly difficult to predict, but it looks like Upper Palaeolithic and Mesolithic hunter-gatherers from Western Europe often had blue eyes, just like Lola. Overall, it looks like Lola’s phenotype—the combination of dark skin, dark hair, and blue eyes—was much more common in prehistoric Europe than it is today.” ref
How Europeans got their lighter skin
“So, why did people in prehistoric Europe look so different from northern Europeans today? The answer to this question lies in a complex interplay between our genes, our changing diets, population movements, and the environment. It has been theorized for some time that lighter skin emerged as an adaptive trait to light poor environments as it allows you to absorb sunlight more effectively, which is essential for the production of vitamin D. However, it was unclear when this happened. Early studies suggested that we first may have evolved lighter skin as our ancestors moved out of Africa and into Europe c. 50,000 years ago, but we now believe that this happened much later in European prehistory. In fact, there is evidence that lighter skin only evolved within the last 5,000 years or so, as a result of genetic admixture from Neolithic farming populations (who carried the skin-lightening variant) and strong selection favoring lighter skin.” ref
Our changing diet also played a part
“In addition, it looks like our changing diets also played a part. During most of European prehistory people relied on wild resources like nuts, game, and fish that are all rich in vitamin D, which is essential to our health. That changed dramatically during the Neolithic when people started to rely on a farmer’s diet that was rich in carbohydrates, but poor in vitamin D. Interestingly, this is exactly the period when we see lighter skin tones evolve in Western Europe and we think that the lack of vitamin D in the diet may have increased the selection pressures favouring lighter skin. All in all, there is solid evidence to suggest that lighter skin tones only evolved in Europe within the last 5,000 years or so, and that people who lived in Europe before then typically had darker skin. It is not that surprising, then, that Lola had darker skin. It simply reflects the fact that she lived at a time when Europeans had not yet evolved their lighter skin.” ref

Groups partially derived from the Ancient North Eurasians
“The ANE lineage is defined by association with the MA-1, or “Mal’ta boy”, remains of 24,000 years ago in central Siberia Mal’ta-Buret’ culture 24,000-15,000 years ago. The Ancient North Eurasians (ANE) samples (Afontova Gora 3, Mal’ta 1, and Yana-RHS) show evidence for minor gene flow from an East Asian-related group (simplified by the Amis, Han, or Tianyuan) but no evidence for ANE-related geneflow into East Asians (Amis, Han, Tianyuan), except the Ainu, of North Japan.” ref
“The ANE lineage is defined by association with the MA-1, or “Mal’ta boy”, remains of 24,000 years ago in central Siberia Mal’ta-Buret’ culture 24,000-15,000 years ago “basal to modern-day Europeans”. Some Ancient North Eurasians also carried East Asian populations, such as Tianyuan Man.” ref
“Bronze-age-steppe Yamnaya and Afanasevo cultures were ANE at around 50% and Eastern Hunter-Gatherer (EHG) at around 75% ANE. Karelia culture: Y-DNA R1a-M417 8,400 years ago, Y-DNA J, 7,200 years ago, and Samara, of Y-haplogroup R1b-P297 7,600 years ago is closely related to ANE from Afontova Gora, 18,000 years ago around the time of blond hair first seen there.” ref
Ancient North Eurasian
“In archaeogenetics, the term Ancient North Eurasian (often abbreviated as ANE) is the name given to an ancestral West Eurasian component that represents descent from the people similar to the Mal’ta–Buret’ culture and populations closely related to them, such as from Afontova Gora and the Yana Rhinoceros Horn Site. Significant ANE ancestry are found in some modern populations, including Europeans and Native Americans.” ref
“The ANE lineage is defined by association with the MA-1, or “Mal’ta boy“, the remains of an individual who lived during the Last Glacial Maximum, 24,000 years ago in central Siberia, Ancient North Eurasians are described as a lineage “which is deeply related to Paleolithic/Mesolithic hunter-gatherers in Europe,” meaning that they diverged from Paleolithic Europeans a long time ago.” ref
“The ANE population has also been described as having been “basal to modern-day Europeans” but not especially related to East Asians, and is suggested to have perhaps originated in Europe or Western Asia or the Eurasian Steppe of Central Asia. However, some samples associated with Ancient North Eurasians also carried ancestry from an ancient East Asian population, such as Tianyuan Man. Sikora et al. (2019) found that the Yana RHS sample (31,600 BP) in Northern Siberia “can be modeled as early West Eurasian with an approximately 22% contribution from early East Asians.” ref
“Populations genetically similar to MA-1 were an important genetic contributor to Native Americans, Europeans, Central Asians, South Asians, and some East Asian groups, in order of significance. Lazaridis et al. (2016:10) note “a cline of ANE ancestry across the east-west extent of Eurasia.” The ancient Bronze-age-steppe Yamnaya and Afanasevo cultures were found to have a noteworthy ANE component at ~50%.” ref
“According to Moreno-Mayar et al. 2018 between 14% and 38% of Native American ancestry may originate from gene flow from the Mal’ta–Buret’ people (ANE). This difference is caused by the penetration of posterior Siberian migrations into the Americas, with the lowest percentages of ANE ancestry found in Eskimos and Alaskan Natives, as these groups are the result of migrations into the Americas roughly 5,000 years ago.” ref
“Estimates for ANE ancestry among first wave Native Americans show higher percentages, such as 42% for those belonging to the Andean region in South America. The other gene flow in Native Americans (the remainder of their ancestry) was of East Asian origin. Gene sequencing of another south-central Siberian people (Afontova Gora-2) dating to approximately 17,000 years ago, revealed similar autosomal genetic signatures to that of Mal’ta boy-1, suggesting that the region was continuously occupied by humans throughout the Last Glacial Maximum.” ref
“The earliest known individual with a genetic mutation associated with blonde hair in modern Europeans is an Ancient North Eurasian female dating to around 16000 BCE from the Afontova Gora 3 site in Siberia. It has been suggested that their mythology may have included a narrative, found in both Indo-European and some Native American fables, in which a dog guards the path to the afterlife.” ref
“Genomic studies also indicate that the ANE component was introduced to Western Europe by people related to the Yamnaya culture, long after the Paleolithic. It is reported in modern-day Europeans (7%–25%), but not of Europeans before the Bronze Age. Additional ANE ancestry is found in European populations through paleolithic interactions with Eastern Hunter-Gatherers, which resulted in populations such as Scandinavian Hunter-Gatherers.” ref
“The Ancient North Eurasians (ANE) split from the ancestors of European peoples somewhere in the Middle East or South-central Asia, and used a northern dispersal route through Central Asia into Northern Asia and Siberia. Genetic analyses show that all ANE samples (Afontova Gora 3, Mal’ta 1, and Yana-RHS) show evidence for minor gene flow from an East Asian-related group (simplified by the Amis, Han, or Tianyuan). In contrast, no evidence for ANE-related geneflow into East Asians (Amis, Han, Tianyuan), except the Ainu, was found.” ref
“Genetic data suggests that the ANE formed during the Terminal Upper-Paleolithic (36+-1,5ka) period from a deeply European-related population, which was once widespread in Northern Eurasia, and from an early East Asian-related group, which migrated northwards into Central Asia and Siberia, merging with this deeply European-related population. These population dynamics and constant northwards geneflow of East Asian-related ancestry would later gave rise to the “Ancestral Native Americans” and Paleosiberians, which replaced the ANE as dominant population of Siberia.” ref
Groups partially derived from the Ancient North Eurasians
“Eastern Hunter-Gatherer (EHG) is a lineage derived predominantly (75%) from ANE. It is represented by two individuals from Karelia, one of Y-haplogroup R1a-M417, dated c. 8.4 kya, the other of Y-haplogroup J, dated c. 7.2 kya; and one individual from Samara, of Y-haplogroup R1b-P297, dated c. 7.6 kya. This lineage is closely related to the ANE sample from Afontova Gora, dated c. 18 kya. After the end of the Last Glacial Maximum, the Western Hunter-Gatherers (WHG) and EHG lineages merged in Eastern Europe, accounting for early presence of ANE-derived ancestry in Mesolithic Europe. Evidence suggests that as Ancient North Eurasians migrated West from Eastern Siberia, they absorbed Western Hunter-Gatherers and other West Eurasian populations as well.” ref
“Caucasian Hunter-Gatherer (CHG) is represented by the Satsurblia individual dated ~13 kya (from the Satsurblia cave in Georgia), and carried 36% ANE-derived admixture. While the rest of their ancestry is derived from the Dzudzuana cave individual dated ~26 kya, which lacked ANE-admixture, Dzudzuana affinity in the Caucasus decreased with the arrival of ANE at ~13 kya Satsurblia.” ref
“Scandinavian Hunter-Gatherer (SHG) is represented by several individuals buried at Motala, Sweden ca. 6000 BC. They were descended from Western Hunter-Gatherers who initially settled Scandinavia from the south, and later populations of EHG who entered Scandinavia from the north through the coast of Norway.” ref
“Iran Neolithic (Iran_N) individuals dated ~8.5 kya carried 50% ANE-derived admixture and 50% Dzudzuana-related admixture, marking them as different from other Near-Eastern and Anatolian Neolithics who didn’t have ANE admixture. Iran Neolithics were later replaced by Iran Chalcolithics, who were a mixture of Iran Neolithic and Near Eastern Levant Neolithic.” ref
“Ancient Beringian/Ancestral Native American are specific archaeogenetic lineages, based on the genome of an infant found at the Upward Sun River site (dubbed USR1), dated to 11,500 years ago. The AB lineage diverged from the Ancestral Native American (ANA) lineage about 20,000 years ago.” ref
“West Siberian Hunter-Gatherer (WSHG) are a specific archaeogenetic lineage, first reported in a genetic study published in Science in September 2019. WSGs were found to be of about 30% EHG ancestry, 50% ANE ancestry, and 20% to 38% East Asian ancestry.” ref
“Western Steppe Herders (WSH) is the name given to a distinct ancestral component that represents descent closely related to the Yamnaya culture of the Pontic–Caspian steppe. This ancestry is often referred to as Yamnaya ancestry or Steppe ancestry.” ref
“Late Upper Paeolithic Lake Baikal – Ust’Kyakhta-3 (UKY) 14,050-13,770 BP were mixture of 30% ANE ancestry and 70% East Asian ancestry.” ref
“Lake Baikal Holocene – Baikal Eneolithic (Baikal_EN) and Baikal Early Bronze Age (Baikal_EBA) derived 6.4% to 20.1% ancestry from ANE, while rest of their ancestry was derived from East Asians. Fofonovo_EN near by Lake Baikal were mixture of 12-17% ANE ancestry and 83-87% East Asian ancestry.” ref
“Hokkaido Jōmon people specifically refers to the Jōmon period population of Hokkaido in northernmost Japan. Though the Jōmon people themselves descended mainly from East Asian lineages, one study found an affinity between Hokkaido Jōmon with the Northern Eurasian Yana sample (an ANE-related group, related to Mal’ta), and suggest as an explanation the possibility of minor Yana gene flow into the Hokkaido Jōmon population (as well as other possibilities). A more recent study by Cooke et al. 2021, confirmed ANE-related geneflow among the Jōmon people, partially ancestral to the Ainu people. ANE ancestry among Jōmon people is estimated at 21%, however, there is a North to South cline within the Japanese archipelago, with the highest amount of ANE ancestry in Hokkaido and Tohoku.” ref

ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref
“A 2016 study found that the global maximum of Ancient North Eurasian (ANE) ancestry occurs in modern-day Kets, Mansi, Native Americans, and Selkups. ANE ancestry has spread throughout Eurasia and the Americas in various migrations since the Upper Paleolithic, and more than half of the world’s population today derives between 5 and 42% of their genomes from the Ancient North Eurasians. Significant ANE ancestry can be found in Native Americans, as well as in regions of northern Europe, South Asia, Central Asia, and Siberia. It has been suggested that their mythology may have featured narratives shared by both Indo-European and some Native American cultures, such as the existence of a metaphysical world tree and a fable in which a dog guards the path to the afterlife.” ref
Ancient Northern East Asian/ later became Ancient Northeast Asian
Ancient Paleo-Siberian
Mal’ta–Buret’ culture (Mal’ta boy MA-1)
The Kolyma Shaitans: Legends and Reality (I only use just a small part)
“A unique “shaitan” burial was discovered on the bank of Omuk-Kuel Lake in the Middle-Kolyma ulus in Yakutia. According to the legends, buried in it are mummified remains of a shaman woman who died during a devastating smallpox epidemics in the 18th c. In an attempt to overcome the deadly disease, the shaman’s relatives used her remains as an emeget fetish. The author believes that these legends reflect the real events of those far-away years. The Arabic word “shaitan” came to the Russian language from Turkic languages. According to Islamic tradition, a shaitan is a genie, an evil spirit, a demon. During Russian colonization and Christianization of Siberia, all sacred things used by the aborigines as fetishes, patron spirits of the family, and the tribe, grew to be called “shaitans.” There are various facts, dating to the 18th and 19th cc., confirming that this word also referred to the mummified remains of outstanding shamans.” ref
“In the 1740s, a member of the Second Kamchatka Expedition Yakov Lindenau wrote, “Meat is scratched off the [shaman’s] bones and the bones are put together to form a skeleton, which is dressed in human’s clothes and worshipped as a deity. The Yukagirs place such dressed bones…in their yurts, their number can sometimes reach 10 or 15. If somebody commits even a minor sacrilege with respect to these bones, he stirs up rancor on the part of the Yukagirs… While traveling and hunting, the Yukagirs carry these bones in their sledges, and moreover, in their best sledges pulled by their best deer. When the Yukagirs are going to undertake something really important, they tell fortune using these skeletons: lift a skeleton up, and if it seems light, it means that their enterprise will have a favorable outcome. The Yukagirs call these skeletons stariks (old men), endow them with their best furs, and sit them on beds covered with deer hides, in a circle, as though they are alive.” (Lindenau, 1983, p. 155)” ref
“In the late 19th c., a famous explorer of aboriginal culture V. I. Jochelson noted the changes that occurred in the ritual in the last century and a half. So, the Yukagirs divided among themselves the shaman’s meat dried in the sun and then put it in separate tents. The dead bodies of killed dogs were left there as well. “After that,” V. I. Jochelson writes, “they would divide the shaman’s bones, dry them and wrap in clothes. The skull was an object of worshipping. It was put on top of a trunk (body) cut out of wood. A caftan and two hats – a winter and a summer one – were sewn for the idol. The caftan was all embroidered. On the skull, a special mask was put, with holes for the eyes and the mouth… The figure was placed in the front corner of the home. Before a meal, a piece of food was thrown into the fire and the idol was held above it. This feeding of the idol… was committed before each meal.” (V. I. Jochelson, 2005, pp. 236—237)” ref
“The idol was kept by the children of the dead shaman. One of them was inducted into the shamanism mysteries while his father was still alive. The idol was carried in a wooden box. Sometimes, in line with the air burial ritual, the box was erected on poles or trees, and the idol was taken out only before hunting or a long journey so that the outcome of the enterprise planned could be predicted. With time, the Yukagirs began using wooden idols as charms. V. I. Jochelson notes that by the late 19th c. the Yukagirs had developed a skeptical attitude towards idols and referred to them as “shaitans.” In this way, under the influence of Christianity, the worshipped ancestor’s spirit turned into its opposite – an evil spirit, a devil, a Satan.” ref
Ancestral Native American, Ancient Beringian
14,000-year-old Ust-Kyakhta-3 (UKY) individual found near Lake Baikal
Chertovy Vorota Cave/Devil’s Gate Cave
Bactria–Margiana Archaeological Complex
32,000-21,000 years ago Yana Culture, at the Yana Woolly Rhinoceros Horn Site in Siberia, with genetic proximity to Ancient North Eurasian populations (Mal’ta and Afontova Gora), but also Ust-Ishim, Sunghir, and to a lesser extent Tianyuan, as well as similarities with the Clovis culture


Paleolithic to Bronze Age Siberians Reveal Connections with First Americans and across Eurasia
“An Upper Paleolithic Siberian shows a deep link with the First Peoples of the Americas. A 10,000-year continuum of Ancient North Eurasian ancestry in the Lake Baikal region. The Neolithic to Bronze Age population formation occurred through prolonged local admixture.” ref
“Modern humans have inhabited the Lake Baikal region since the Upper Paleolithic, though the precise history of its peoples over this long time span is still largely unknown. Here, we report genome-wide data from 19 Upper Paleolithic to Early Bronze Age individuals from this Siberian region. An Upper Paleolithic genome shows a direct link with the First Americans by sharing the admixed ancestry that gave rise to all non-Arctic Native Americans. We also demonstrate the formation of Early Neolithic and Bronze Age Baikal populations as the result of prolonged admixture throughout the eighth to sixth millennium years ago. Moreover, we detect genetic interactions with western Eurasian steppe populations and reconstruct Yersinia pestis genomes from two Early Bronze Age individuals without western Eurasian ancestry. Overall, our study demonstrates the most deeply divergent connection between Upper Paleolithic Siberians and the First Americans and reveals human and pathogen mobility across Eurasia during the Bronze Age.” ref
“The Lake Baikal region in Siberia has been inhabited by modern humans since the Upper Paleolithic and has a rich archaeological record. In the past 5 years, ancient genomic studies have revealed multiple genetic turnovers and admixture events in this region. The 24,000-year-old individual (MA1) from the Mal’ta site represents an ancestry referred to as “Ancient North Eurasian (ANE),” which was widespread across Siberia during the Paleolithic and that contributed to the genetic profile of a vast number of present-day Eurasian populations as well as Native Americans. ANE ancestry was suggested to have been largely replaced in the Lake Baikal region during the Early Neolithic by a gene pool related to present-day northeast Asians, with a limited resurgence of ANE ancestry by the Early Bronze Age.” ref
“Siberia has also been proposed as a source for multiple waves of dispersals into the Americas, the first of which was shown to be driven by a founding population estimated to have formed around 25,000–20,000 years ago. The so-called Ancient Beringian ancestry represented by a 11,500-year-old Alaskan individual (USR1) was shown to be part of this founding population, estimated to have split from other Native Americans around 23,000 years ago. In addition, the recently published 9,800-year-old Kolyma genome from northeastern Siberia was suggested to represent the closest relative to Native American populations outside of the Americas. Moreover, the Paleo-Eskimo ancestry represented by a 4,000-year-old Saqqaq individual from Greenland was also estimated to have split from northeastern Siberian groups and migrated to Arctic America around 6,000–5,000 years ago. Although these waves of migration are generally linked to ancient Siberian populations, their origins in the context of the Siberian genetic history remain poorly understood. Further studies of the Siberian population history using ancient genomes are, therefore, critical for the better understanding of the formation of Native American populations.” ref
“Furthermore, the Neolithic to Bronze Age transition in Eurasia was marked by complex cultural and genetic changes facilitated by extensive population movements, though their impact in the Lake Baikal region is still unclear. Looking to the west, the Early Bronze Age groups from the Pontic-Caspian steppe associated with the Yamnaya complex spread both east and west along with their distinct genetic profile often referred to as “Steppe ancestry”. The eastward expansion of this group is considered to be associated with the Early Bronze Age Afanasievo culture. However, the later Middle Bronze Age Okunevo-related population from the central steppe, as well as the Late Bronze Age Khövsgöl-related population from the eastern steppe, harbor only a limited proportion of Steppe ancestry. Therefore, the effect of steppe migrations in eastern Eurasia, particularly the interactions of Bronze Age Baikal hunter-gatherers with the contemporaneous and geographically proximal Afanasievo population, is still largely unexplored.” ref
“In this study, researchers report 19 newly sequenced ancient hunter-gatherers from the Lake Baikal and its surrounding regions, spanning from the Upper Paleolithic to the Early Bronze Age. Their analyses alongside published data reveal the most deeply divergent ancestry that link Upper Paleolithic Siberians and the First Peoples of the Americas, and more clearly delineate the complex transition between Early Neolithic and Early Bronze Age populations in the Lake Baikal region. We also provide both human and pathogen genomic evidence demonstrating the influence of western Eurasian steppe populations in this region during the Early Bronze Age and discuss the genetic contribution of Lake Baikal hunter-gatherers to Siberian populations through time.” ref
“Most of the Lake Baikal individuals occupied the space on a “ANE-NEA” cline running between “Northeast Asian” (NEA) ancestry represented by Neolithic hunter-gathers from the Devil’s Gate in the Russian Far East, and the ANE ancestry represented by Upper Paleolithic Siberian individuals MA1, AfontovaGora 2 (AG2), and AfontovaGora 3 (AG3), which was first described by. Our newly sequenced Upper Paleolithic genome from the Ust-Kyakhta-3 site (UKY) just south to the Lake Baikal is placed close to the Mesolithic northeastern Siberian Kolyma individual and is shifted toward Native American populations compared to the rest of the ancient Baikal individuals along PC2. All four Early Neolithic individuals cluster with published Early Neolithic groups from the same region (Shamanka_EN, Lokomotiv_EN, UstBelaya_EN) designated as the “Baikal_EN” population. The LNBA individuals were divided into four groups. The major “Baikal_LNBA” group included 10 individuals and clustered with published Late Neolithic to Bronze Age Baikal populations (Shamanka_EBA, Kurma_EBA, UstIda_EBA, UstIda_LN, UstBelaya_BA).” ref
“These individuals were positioned in PCA closer to ANE-related individuals compared with the Early Neolithic individuals from the same region, as well as closer to the Paleo-Eskimo Saqqaq individual. Another two individuals (GLZ001 and GLZ002) from the Glazkovskoe predmestie site, unlike the third individual from the same archaeological site (GLZ003), seemed shifted from the main cluster and showed closer genetic affinity to the Devil’s Gate and Early Neolithic Baikal individuals. One of the six individuals from the Kachug site (KPT005) was substantially displaced from the Baikal_LNBA group toward western Eurasians along PC1, not along the ANE-NEA cline but toward later Bronze Age populations, suggesting a potential introgression of the Steppe-related ancestry. Finally, an Early Bronze Age individual (BZK002) from the Bazaikha site in the Yenisei River region further to the west of the Lake Baikal was significantly displaced toward ANE-related individuals and located close to published Bronze Age individuals associated to the Okunevo culture.” ref
“Population clustering with ADMIXTURE based on worldwide populations also showed a similar clustering pattern. When selecting a K value of 16, the published and newly sequenced individuals belonging to main Early Neolithic to Bronze Age Baikal groups all showed genetic profiles composed of a mixture of three major components that were mostly enriched in ANE-related individuals, northeast Asians, and central Siberians represented by the Uralic-speaking Nganasan population. The ANE and central Siberian ancestries were both of higher proportion in most LNBA Baikal individuals than in the Early Neolithic ones, while GLZ001 and GLZ002 showed higher NEA ancestry, similar to the Early Neolithic population. The BZK002 individual presented a profile similar to the published Okunevo group, with a much larger ANE component compared to other Lake Baikal individuals. The KPT005 individual also displayed a substantial contribution derived from European “Western Hunter-Gatherer” (WHG) ancestry, likely acquired through gene flow from the west.” ref
“Researchers estimated the runs of homozygosity (ROH) of selected individuals together with published Baikal individuals and did not identify an inbreeding signal in any individual. The Kolyma individual showed significantly more ROH compared with other individuals, suggesting a smaller population size in Mesolithic northeastern Siberia. The sharing of identity-by-descent (IBD) segments between individuals suggested a close relationship between UKY and Kolyma, supporting our analyses based on genome-wide SNP data, and also revealed that Early Neolithic and LNBA Baikal individuals shared genetic affinity with each other as well as with the older UKY and Kolyma genomes.” ref

World’s oldest known fort was constructed by hunter-gatherers 8,000 years ago in Siberia
“The fact that this Stone Age fort was built by hunter-gatherers is transforming our understanding of ancient human societies. Hunter-gatherers built the oldest known fort in the world about 8,000 years ago in Siberia, a new study finds. Archaeologists have long associated fortresses with permanent agricultural settlements. However, this cluster of fortified structures reveals that prehistoric groups were constructing protective edifices much earlier than originally thought.” ref
“These hunter-gatherers “defy conventional stereotypes that depict such societies as basic and nomadic, unveiling their capacity to construct intricate structures,” study co-author Tanja Schreiber, an archaeologist at Free University of Berlin, told Live Science in an email. Located along the Amnya River in western Siberia, remains of the Amnya fort include roughly 20 pit-house depressions scattered across the site, which is divided into two sections: Amnya I and Amnya II. Radiocarbon dating confirmed that the settlement was first inhabited during the Mesolithic, or Middle Stone Age, according to the study. When constructed, each pit house would have been protected by earthen walls and wooden palisades — two construction elements that suggest “advanced agricultural and defensive capabilities” by the inhabitants, the archaeologists said in a statement.” ref
“One of the Amnya fort’s most astonishing aspects is the discovery that approximately 8,000 years ago, hunter-gatherers in the Siberian Taiga built intricate defense structures,” Schreiber said. “This challenges traditional assumptions that monumental constructions were solely the work of agricultural communities.” It’s unknown what triggered the need for these fortified structures in the first place, but the strategic location overlooking the river would have not only been an ideal lookout point for potential threats but also allowed hunter-gatherers to keep tabs on their fishing and hunting grounds, the researchers noted.” ref

“The Samara culture was an Eneolithic (Copper Age) culture that flourished around the turn of the 5th millennium BCE, at the Samara Bend of the Volga River (modern Russia). The Samara culture is regarded as related to contemporaneous or subsequent prehistoric cultures of the Pontic–Caspian steppe, such as the Khvalynsk, Repin, and Yamna (or Yamnaya) cultures.” ref
“Genetic analyses of a male buried at Lebyazhinka, radiocarbon dated to 5640-5555 BCE, found that he belonged to a population often referred to as “Samara hunter-gatherers”, a group closely associated with Eastern Hunter-Gatherers. The male sample carried Y-haplogroup R1b1a1a and mitochondrial haplogroup U5a1d.” ref
“Pottery consists mainly of egg-shaped beakers with pronounced rims. They were not able to stand on a flat surface, suggesting that some method of supporting or carrying must have been in use, perhaps basketry or slings, for which the rims would have been a useful point of support. The carrier slung the pots over the shoulder or onto an animal. The decoration consists of circumferential motifs: lines, bands, zig-zags, or wavy lines, incised, stabbed, or impressed with a comb. These patterns are best understood when seen from the top. They appear then to be a solar motif, with the mouth of the pot as the sun. Later developments of this theme show that in fact the sun is being represented.” ref
“The culture is characterized by the remains of animal sacrifice, which occur over most of the sites. There is no indisputable evidence of riding, but there were horse burials, the earliest in the Old World. Typically the head and hooves of cattle, sheep, and horses are placed in shallow bowls over the human grave, smothered with ochre. Some have seen the beginning of the horse sacrifice in these remains, but this interpretation has not been more definitely substantiated. We know that the Indo-Europeans sacrificed both animals and people, like many other cultures.” ref
“The graves found are shallow pits for single individuals, but two or three individuals might be placed there. Some of the graves are covered with a stone cairn or a low earthen mound, the very first predecessor of the kurgan. The later, fully developed kurgan was a hill on which the deceased chief might ascend to the sky god, but whether these early mounds had that significance is doubtful.” ref
“Grave offerings included ornaments depicting horses. The graves also had an overburden of horse remains; it cannot yet be determined decisively if these horses were domesticated and ridden or not, but they were certainly used as a meat-animal. Most controversial are bone plaques of horses or double oxen heads, which were pierced. The graves yield well-made daggers of flint and bone, placed at the arm or head of the deceased, one in the grave of a small boy. Weapons in the graves of children are common later. Other weapons are bone spearheads and flint arrowheads. Other carved bone figurines and pendants were found in the graves.” ref
“The Yamnaya culture or the Yamna culture, also known as the Pit Grave culture or Ochre Grave culture, was a late Copper Age to early Bronze Age archaeological culture of the region between the Southern Bug, Dniester, and Ural rivers (the Pontic–Caspian steppe), dating to 3300–2600 BCE or around 5,300 to 4,600 years ago. It was discovered by Vasily Gorodtsov following his archaeological excavations near the Donets River in 1901–1903. Its name derives from its characteristic burial tradition: Я́мная (romanization: yamnaya) is a Russian adjective that means ‘related to pits (yama)’, as these people used to bury their dead in tumuli (kurgans) containing simple pit chambers.” ref
“The Yamnaya economy was based upon animal husbandry, fishing, and foraging, and the manufacture of ceramics, tools, and weapons. The people of the Yamnaya culture lived primarily as nomads, with a chiefdom system and wheeled carts and wagons that allowed them to manage large herds. They are also closely connected to Final Neolithic cultures, which later spread throughout Europe and Central Asia, especially the Corded Ware people and the Bell Beaker culture, as well as the peoples of the Sintashta, Andronovo, and Srubnaya cultures.” ref
“Back migration from Corded Ware also contributed to Sintashta and Andronovo. In these groups, several aspects of the Yamnaya culture are present. Yamnaya material culture was very similar to the Afanasevo culture of South Siberia, and the populations of both cultures are genetically indistinguishable. This suggests that the Afanasevo culture may have originated from the migration of Yamnaya groups to the Altai region or, alternatively, that both cultures developed from an earlier shared cultural source.” ref
“Genetic studies have suggested that the people of the Yamnaya culture can be modelled as a genetic admixture between a population related to Eastern European Hunter-Gatherers (EHG) and people related to hunter-gatherers from the Caucasus (CHG) in roughly equal proportions, an ancestral component which is often named “Steppe ancestry”, with additional admixture from Anatolian, Levantine, or Early European farmers. Genetic studies also indicate that populations associated with the Corded Ware, Bell Beaker, Sintashta, and Andronovo cultures derived large parts of their ancestry from the Yamnaya or a closely related population.” ref
“The origin of the Yamnaya culture continues to be debated, with proposals for its origins pointing to both the Khvalynsk and Sredny Stog cultures. The Khvalynsk culture (4700–3800 BCE) (middle Volga) and the Don-based Repin culture (c. 3950–3300 BCE) in the eastern Pontic-Caspian steppe, and the closely related Sredny Stog culture (c. 4500–3500 BCE) in the western Pontic-Caspian steppe, preceded the Yamnaya culture (3300–2500 BCE). The Yamnaya culture was succeeded in its western range by the Catacomb culture (2800–2200 BCE); in the east, by the Poltavka culture (2700–2100 BCE) at the middle Volga. These two cultures were followed by the Srubnaya culture (18th–12th century BCE).” ref
“Further efforts to pinpoint the location came from Anthony (2007), who suggested that the Yamnaya culture (3300–2600 BCE) originated in the Don–Volga area at c. 3400 BCE, preceded by the middle Volga-based Khvalynsk culture and the Don-based Repin culture (c. 3950–3300 BCE), arguing that late pottery from these two cultures can barely be distinguished from early Yamnaya pottery. Earlier continuity from eneolithic but largely hunter-gatherer Samara culture and influences from the more agricultural Dnieper–Donets II are apparent.” ref
“He argues that the early Yamnaya horizon spread quickly across the Pontic–Caspian steppes between c. 3400 and 3200 BCE:
The spread of the Yamnaya horizon was the material expression of the spread of late Proto-Indo-European across the Pontic–Caspian steppes.
[…] The Yamnaya horizon is the visible archaeological expression of a social adjustment to high mobility – the invention of the political infrastructure to manage larger herds from mobile homes based in the steppes.” ref
“Alternatively, Parpola (2015) relates both the Corded ware culture and the Yamnaya culture to the late Trypillia (Tripolye) culture. He hypothesizes that “the Tripolye culture was taken over by PIE speakers by c. 4000 BCE,” and that in its final phase the Trypillian culture expanded to the steppes, morphing into various regional cultures which fused with the late Sredny Stog (Serednii Stih) pastoralist cultures, which, he suggests, gave rise to the Yamnaya culture. Dmytro Telegin viewed Sredny Stog and Yamna as one cultural continuum and considered Sredny Stog to be the genetic foundation of the Yamna.” ref
“The Yamnaya culture was nomadic or semi-nomadic, with some agriculture practiced near rivers, and a few fortified sites, the largest of which is Mikhaylivka. Characteristic for the culture are the burials in pit graves under kurgans (tumuli), often accompanied by animal offerings. Some graves contain large anthropomorphic stelae, with carved human heads, arms, hands, belts, and weapons. The dead bodies were placed in a supine position with bent knees and covered in ochre. Some kurgans contained “stratified sequences of graves.” ref
“Kurgan burials may have been rare, and were perhaps reserved for special adults, who were predominantly, but not necessarily, male. Status and gender are marked by grave goods and position, and in some areas, elite individuals are buried with complete wooden wagons. Grave goods are more common in eastern Yamnaya burials, which are also characterized by a higher proportion of male burials and more male-centred rituals than western areas.” ref
“The Yamnaya culture had and used two-wheeled carts and four-wheeled wagons, which are thought to have been oxen-drawn at this time, and there is evidence that they rode horses. For instance, several Yamnaya skeletons exhibit specific characteristics in their bone morphology that may have been caused by long-term horseriding. Metallurgists and other craftsmen are given a special status in Yamnaya society, and metal objects are sometimes found in large quantities in elite graves.” ref
“New metalworking technologies and weapon designs are used. Stable isotope ratios of Yamna individuals from the Dnipro Valley suggest the Yamnaya diet was terrestrial protein based with insignificant contribution from freshwater or aquatic resources. Anthony speculates that the Yamnaya ate meat, milk, yogurt, cheese, and soups made from seeds and wild vegetables, and probably consumed mead.” ref
“Mallory and Adams suggest that Yamnaya society may have had a tripartite structure of three differentiated social classes, although the evidence available does not demonstrate the existence of specific classes such as priests, warriors, and farmers.” ref
“According to Jones et al. (2015) and Haak et al. (2015), autosomal tests indicate that the Yamnaya people were the result of a genetic admixture between two different hunter-gatherer populations: distinctive “Eastern Hunter-Gatherers” (EHG), from Eastern Europe, with high affinity to the Mal’ta–Buret’ culture or other, closely related people from Siberia and a population of “Caucasus hunter-gatherers” (CHG) who probably arrived from the Caucasus or Iran. Each of those two populations contributed about half the Yamnaya DNA. This admixture is referred to in archaeogenetics as Western Steppe Herder (WSH) ancestry.” ref
“Admixture between EHGs and CHGs is believed to have occurred on the eastern Pontic-Caspian steppe starting around 5,000 BCE, while admixture with Early European Farmers (EEF) happened in the southern parts of the Pontic-Caspian steppe sometime later. More recent genetic studies have found that the Yamnaya were a mixture of EHGs, CHGs, and to a lesser degree Anatolian farmers and Levantine farmers, but not EEFs from Europe due to lack of WHG DNA in the Yamnaya. This occurred in two distinct admixture events from West Asia into the Pontic-Caspian steppe.” ref
“Haplogroup R1b, specifically the Z2103 subclade of R1b-L23, is the most common Y-DNA haplogroup found among the Yamnaya specimens. This haplogroup is rare in Western Europe and mainly exists in Southeastern Europe today. Additionally, a minority are found to belong to haplogroup I2. They are found to belong to a wider variety of West Eurasian mtDNA haplogroups, including U, T, and haplogroups associated with Caucasus Hunter-Gatherers and Early European Farmers. A small but significant number of Yamnaya kurgan specimens from Northern Ukraine carried the East Asian mtDNA haplogroup C4.” ref
“In 2014, a study discovered a new mtDNA subclade C1f from the remains of 3 people found in north-western Russia and dated to 7,500 years ago. The subclades C1b, C1c, C1d, and C4c are found in the first people of the Americas. C1a is found only in Asia.” ref
“C4 – Upper Palaeolithic (14050 – 13770 years ago) Ust-Kyakhta (Buryatia), Late Neolithic-Bronze Age Irkutsk Oblast, Late Neolithic-Iron Age Yakutia, Tubalar (Ederbes), Todzhin (Toora-Hem, Iiy, Adir-Kezhig), Yukaghir (Andrushkino), Yukaghir/Chuvan (Markovo), Russian, Myanmar
-
- C4a’b’c – Irkutsk Oblast (6815 years ago), India (Jenu Kuruba)
- C4a – China (Guangdong, Han from Beijing)
- C4a1 – Mongol from Chifeng and Hulunbuir, Tashkurgan (Kyrgyz, Sarikoli, Wakhi), Czech Republic, Denmark
- C4a1a – Korea, China, Uyghur, Buryat (South Siberia), Denmark, Sweden, France, Scotland, Canada
- C4a1a1
- C-T195C! – Ireland, Scotland, England, USA, Hungary (Szeged region), Poland, Belarus, Russia (Russian, Buryat), Turkey, Pakistan (Hazara), India (Jammu and Kashmir), China (Bargut and Mongol in Inner Mongolia, etc.), Korea
- C4a1a2 – China
- C4a1a2a – China (Han from Ili, Han from Henan, etc.)
- C4a1a2b
- C4a1a2b1 – China
- C4a1a2b2 – Uyghur
- C4a1a3 – Bronze Age Irkutsk Oblast (Ust’-Belaya, Khaptsagai, Silinskij, Chastaja Padi), Russian (Kemerovo Oblast), Koryak, Yukaghir, Yakut, Evenk (Nyukzha, Chumikan, Nelkan/Dzhigda), Even (Sakkyryyr, Sebjan, Tompo, Markovo, Kamchatka), Udinsk Buryat (Kushun), Todzhin (Toora-Hem, Adir-Kezhig), Altai Kizhi, Iran (Qashqai), Sweden
- C4a1a3a – Yakut, Buryat (Buryat Republic, Irkutsk Oblast), Bargut, Nentsi
- C4a1a3a1 – Yakut, Nganasan (Vadei of Taimyr Peninsula)
- C4a1a3a1a – Evenk (Taimyr, Stony Tunguska)
- C4a1a3a1b – Tofalar
- C4a1a3a1 – Yakut, Nganasan (Vadei of Taimyr Peninsula)
- C4a1a3b – Bargut, Uyghur
- C4a1a3b1 – Chelkan, Tubalar
- C4a1a3c – Evenk (Taimyr Peninsula, Stony Tunguska)
- C4a1a3d – Yakut
- C4a1a3a – Yakut, Buryat (Buryat Republic, Irkutsk Oblast), Bargut, Nentsi
- C4a1a4 – Buryat, Kazakhstan
- C4a1a4a – Evenk (Okhotsk region), Shor
- C4a1a2 – China
- C4a1a5 – Teleut, Ladakh
- C4a1a6
- C4a1a6a – Russia (Bashkortostan, Khamnigan), Kyrgyzstan (Kyrgyz), Inner Mongolia (Bargut, Buryat)
- C4a1a6b – Buryat (South Siberia, Inner Mongolia), Uyghur
- C4a1a7 – Denmark
- C4a1b – China, Thailand (Palaung)
- C4a1c – Russia (Bashkortostan, Adygei), Iran (Azerbaijanian), China (Xibo, Mongol from Tianjin)
- C4a1a – Korea, China, Uyghur, Buryat (South Siberia), Denmark, Sweden, France, Scotland, Canada
- C4a2
- C4a2a – Yakut, Evenk (Chumikan)
- C4a2a1 – Bronze Age (2275 – 2040 cal BCE or around 4,275 to 4,040 years ago) Irkutsk Oblast (specimen irk076 from burial 3 at the Shamanka 2 site, South Baikal), Shor, Chelkan, Teleut, Altai Kizhi, Yakut, Kazakh, Ket, Evenk (Stony Tunguska, Taimyr), Buryat (Irkutsk Oblast, Inner Mongolia), China, Korea
- C4a2a1a – Yukaghir, Yakut, Evenk (Nyukzha, Iyengra, Nelkan/Dzhigda), Even (Tompo)
- C4a2a1b – Evenk (Nyukzha), Yakut
- C4a2a1b1 – Evenk (Nyukzha)
- C4a2a1c – China (Zhejiang, Uyghurs), Buryat, Todzhin (Iiy), Karanogay (Dagestan)
- C4a2a1c1 – Tofalar (Alygdzher, Nerkha, V. Gutara), Khamnigan
- C4a2a1c2 – Uyghurs
- C4a2a1d – Uyghurs
- C4a2a1d1 – Udinsk Buryat (Kushun), Tofalar (V. Gutara), Evenk (Central Siberia)
- C4a2a1d2 – Evenk (Nelkan/Dzhigda), Evenk/Nivkh (Val)
- C4a2a1e – Bargut (Inner Mongolia), Buryat (Irkutsk Oblast)
- C4a2a1f – Buryat (South Siberia, Irkutsk Oblast)
- C4a2a1g – Ket
- C4a2a1 – Bronze Age (2275 – 2040 cal BCE or around 4,275 to 4,040 years ago) Irkutsk Oblast (specimen irk076 from burial 3 at the Shamanka 2 site, South Baikal), Shor, Chelkan, Teleut, Altai Kizhi, Yakut, Kazakh, Ket, Evenk (Stony Tunguska, Taimyr), Buryat (Irkutsk Oblast, Inner Mongolia), China, Korea
- C4a2b – Tibet, Korea
- C4a2b1 – Wancho
- C4a2b2 – China (Han from Beijing)
- C4a2b2a – Tibet (Sherpa)
- C4a2c – Bargut (Inner Mongolia)
- C4a2c1 – India (Jenu Kuruba)
- C4a2c2 – Lepcha
- C4a2c2a – Ladakh
- C4a2a – Yakut, Evenk (Chumikan)
- C4a1 – Mongol from Chifeng and Hulunbuir, Tashkurgan (Kyrgyz, Sarikoli, Wakhi), Czech Republic, Denmark
- C4b – Mongol from Jilin and Hulunbuir, Yukaghir, Altai Kizhi, Ukraine, Slovakia
- C4c – Ijka
- C4c1 – Sioux (Carson County of South Dakota), Shuswap, Canada, USA, France, Spain
- C4c1a – Cherokee (Flint District of Oklahoma)
- C4c1b – Chippewa (Trempealeau in Wisconsin), Ottawa or Chippewa (Sault Saint Marie, Chippewa County, Michigan), Canada
- C4c2 – Métis (Red River, Manitoba), USA
- C4c1 – Sioux (Carson County of South Dakota), Shuswap, Canada, USA, France, Spain
- C4a – China (Guangdong, Han from Beijing)
- C4-T152C! – Russia (Bashkortostan), England
- C4-T152C!-A12780G – Uyghur
- C4d – Turkey, Tibet (Chamdo, Nyingchi, Shannan, Lhoba), Thailand (Khon Mueang from Chiang Mai Province), Han from Beijing, Mongol from Tongliao
- C4-T152C!-T4742C – Altai Republic (ancient DNA), Uyghur
- C4-T152C!-T4742C-T16093C – Kyrgyz (Kyrgyzstan), Tibet (Nyingchi)
- C4-T152C!-T4742C-T8602C – Sarikoli (Tashkurgan), Burusho (Pakistan)
- C4-T152C!-T4742C-T8602C-G11176A – Pamiri (Gorno-Badakhshan Autonomous Region of Tajikistan)
- C4e – Teleut, Shor” ref
- C4-T152C!-A12780G – Uyghur
- C4a’b’c – Irkutsk Oblast (6815 years ago), India (Jenu Kuruba)
“People of the Yamnaya culture are believed to have had mostly brown eye colour, light to intermediate skin, and brown hair colour, with some variation.” ref
“Some Yamnaya individuals are believed to have carried a mutation to the KITLG gene associated with blond hair, as several individuals with Steppe ancestry are later found to carry this mutation. The Ancient North Eurasian Afontova Gora group, who contributed significant ancestry to Western Steppe Herders, are believed to be the source of this mutation. A study in 2015 found that Yamnaya had the highest ever calculated genetic selection for height of any of the ancient populations tested. It has been hypothesized that an allele associated with lactase persistence (conferring lactose tolerance into adulthood) was brought to Europe from the steppe by Yamnaya-related migrations.” ref
“A 2022 study by Lazaridis et al. found that the typical phenotype among the Yamnaya population was brown eyes, brown hair, and intermediate skin colour. None of their Yamnaya samples were predicted to have either blue eyes or blond hair, in contrast with later Steppe groups in Russia and Central Asia, as well as the Bell Beaker culture in Europe, who did carry these phenotypes in high proportions.” ref
“The geneticist David Reich has argued that the genetic data supports the likelihood that the people of the Yamnaya culture were a “single, genetically coherent group” who were responsible for spreading many Indo-European languages. Reich’s group recently suggested that the source of Anatolian and Indo-European subfamilies of the Proto-Indo-European (PIE) language may have been in west Asia and the Yamna were responsible for the dissemination of the latter. Reich also argues that the genetic evidence shows that Yamnaya society was an oligarchy dominated by a small number of elite males.” ref
“The genetic evidence for the extent of the role of the Yamnaya culture in the spread of Indo-European languages has been questioned by Russian archaeologist Leo Klejn and Balanovsky et al., who note a lack of male haplogroup continuity between the people of the Yamnaya culture and the contemporary populations of Europe. Klejn has also suggested that the autosomal evidence does not support a Yamnaya migration, arguing that Western Steppe Herder ancestry in both contemporary and Bronze Age samples is lowest around the Danube in Hungary, near the western limits of the Yamnaya culture, and highest in Northern Europe, which Klejn argues is the opposite of what would be expected if the geneticists’ hypothesis is correct.” ref
Yamnaya culture and the Proto-Indo-Europeans (PIE) Language
“Marija Gimbutas identified the Yamnaya culture with the late Proto-Indo-Europeans (PIE) in her Kurgan hypothesis. In the view of David Anthony, the Pontic-Caspian steppe is the strongest candidate for the Urheimat (original homeland) of the Proto-Indo-European language, citing evidence from linguistics and genetics which suggests that the Yamnaya culture may be the homeland of the Indo-European languages, with the possible exception of the Anatolian languages. On the other hand, Colin Renfrew has argued for a Near Eastern origin of the earliest Indo-European speakers.” ref
“According to David W. Anthony, the genetic evidence suggests that the leading clans of the Yamnaya were of EHG (Eastern European hunter-gatherer) and WHG (Western European hunter-gatherer) paternal origin and implies that the Indo-European languages were the result of “a dominant language spoken by EHGs that absorbed Caucasus-like elements in phonology, morphology, and lexicon.” It has also been suggested that the PIE language evolved through trade interactions in the circum-Pontic area in the 4th millennium BCE, mediated by the Yamna predecessors in the North Pontic steppe.” ref
“Guus Kroonen et al. 2022 found that the “basal Indo-European stage”, also known as Indo-Anatolian or Pre-Proto-Indo-European language, largely but not totally, lacked agricultural-related vocabulary, and only the later “core Indo-European languages” saw an increase in agriculture-associated words. According to them, this fits a homeland of early core Indo-European within the westernmost Yamnaya horizon, around and west of the Dnieper, while its basal stage, Indo-Anatolian, may have originated in the Sredny Stog culture, as opposed to the eastern Yamnaya horizon.” ref
“The Corded Ware culture may have acted as major source for the spread of later Indo-European languages, including Indo-Iranian, while Tocharian languages may have been mediated via the Catacomb culture. They also argue that this new data contradicts a possible earlier origin of Pre-Proto-Indo-European among agricultural societies South of the Caucasus, rather “this may support a scenario of linguistic continuity of local non-mobile herders in the Lower Dnieper region and their genetic persistence after their integration into the successive and expansive Yamnaya horizon”. Furthermore the authors mention that this scenario can explain the difference in paternal haplogroup frequency between the Yamnaya and Corded Ware cultures, while both sharing similar autosomal DNA ancestry.” ref

ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref
Early Russian Pottery in Cisbaikal Kitoi culture 7,500 years ago, Samara culture 7,000 years ago, and Yamnaya culture 5,600–4,600 years ago, as well as Proto-Indo-European emergence
“The area east of Lake Baikal in Siberia is one of the few regions in Eurasia where pottery was already used during the Late Pleistocene and Early Holocene. Such early pottery complexes were identified in Ust’-Karenga XII, Studenoye 1, Ust’-Menza 1, and Ust’-Khyakhta 3, dated at about 12-000-11,000 years ago. While around 20,000 years ago East Asian hunter-gatherers were already making ceramic pots. (It seems to Damien) that ceramics spread continually from the earliest centers in China, then Japan, and next the Russian Far East, lastly towards the west, all the way to Europe.” ref
(ANA) Ancient Northeast Asian ancestry today
“Genetically, ANA/Amur ancestry peaks among modern Tungusic, Mongolic, and

“Abstract: Occupied between ~10,300 and 9300 years ago, the Pre-Pottery Neolithic site of Aşıklı Höyük in Central Anatolia went through early phases of sheep domestication. Analysis of 629 mitochondrial genomes from this and numerous sites in Anatolia, southwest Asia, Europe, and Africa produced a phylogenetic tree with excessive coalescences (nodes) around the Neolithic, a potential signature of a domestication bottleneck. This is consistent with archeological evidence of sheep management at Aşıklı Höyük which transitioned from residential stabling to open pasturing over a millennium of site occupation. However, unexpectedly, we detected high genetic diversity throughout Aşıklı Höyük’s occupation rather than a bottleneck. Instead, we detected a tenfold demographic bottleneck later in the Neolithic, which caused the fixation of mitochondrial haplogroup B in southwestern Anatolia. The mitochondrial genetic makeup that emerged was carried from the core region of early Neolithic sheep management into Europe and dominates the matrilineal diversity of both its ancient and the billion-strong modern sheep populations.” ref
“The establishment of Neolithic sedentary societies in southwest Asia was associated with the development of farming practices between 10,000 and 12,000 years ago. Those practices included the cultivation of cereals and legumes and the management of sheep, goats, cattle, and pigs, which ultimately resulted in their domestication. Crop-livestock subsistence strategies started gaining ground around 10,500 years ago in the northern “Fertile Crescent,” and by 9,500 years ago, this mode of subsistence had replaced the foraging lifestyle in parts of southwest Asia and Cyprus. Archaeobotanical and zooarcheological analyses showed that these millennia-long practices of crop cultivation and ungulate management led to phenotypic changes in both plants and animals. To understand these changes, it is often necessary to integrate the evidence of multiple sites and millennia. However, only a few Pre-Pottery Neolithic sites preserved a sufficiently long occupation history and representative faunal assemblages to track morphological, biometric, and demographic changes related to early livestock management at a single location. The list of such exceptional sites in Anatolia includes Çayönü, Cafer Höyük, and Nevalı Çori in Southeastern Anatolia and Aşıklı Höyük in Central Anatolia.” ref
“Aşıklı Höyük is situated on the bank of the Melendiz River. Here, large numbers of caprine bones (i.e., sheep and goats) have been excavated from occupational phases spanning over a thousand years, between ~10,300 and 9,300 years ago. The importance of small livestock management at the site was such that, during the thousand-years occupation, the composition of animal remains identified as sheep and goat increased from ~50% to 87%. Analyses of this extraordinary assemblage provided a unique glimpse into early strategies of sheep management. This includes mortality curves that are indicative of the culling of juvenile males, which in turn reflects exploitation for meat, and spatial patterns of skeletal distribution that imply that slaughtering took place near the living quarters. Further analyses of intra-articular joint pathologies suggested restricted mobility close to the village, including residential stabling, which led to the accumulation of dung and urine salts in the sediments.” ref
“By 9,700 years ago, however, sheep management strategies apparently shifted toward extensive herding. Evidence for this includes a decrease in urine salt and dung concentrations in residential areas, an increase in carcass size (table S2), an improvement in joint health implying greater mobility, and shifts in phytolith and stable isotope profiles that imply more extensive grazing. Together, the evidence obtained at Aşıklı Höyük demonstrates that sheep management in early Neolithic communities was a dynamic process of learning by doing. Although management strategies at Aşıklı Höyük likely affected the phenotype of sheep populations, it is not clear whether they initiated evolutionary changes that ultimately led to the strong genetic differentiation between wild and domestic populations that we observe today.” ref
“A common assumption is that capture and spatial isolation of a subset of a wild population induced a “domestication bottleneck,” provoking the general reduction of genetic diversity evident in modern domestic sheep populations. Here, to address whether the initial management of sheep at Aşıklı Höyük caused shifts in their genetic makeup, we analyzed 629 whole mitogenomes sourced from 15 countries, including 62 from Aşıklı Höyük, spanning a period of over 10,000 years. This allowed us to infer the mitochondrial phylogeography and maternal demographic history of Anatolian and European sheep, and the contribution of the Aşıklı Höyük community to the formation of the Neolithic package dispersing across north and southwestern Anatolia between 10,000 and 8,000 years ago, and subsequently into Europe.” ref
“When we tested the temporal changes in haplogroup frequencies, we found, in general, significant differences between, but not within, groups. The western group included Africa, Europe (modern and ancient), and Neolithic southwestern Anatolia, while the eastern group included Neolithic Aşıklı Höyük, Chalcolithic Güvercinkayası, Caucasus, Levant, and eastern Asia. This pattern was true when the tests were performed with effective population sizes (Ne) between 104 and 106 (we refrained from writing P values here because of the numerous combinations of Ne used in these analyses; see table S6 for the exact values and fig. S2 for a graphical summary of these results). The whole pattern of significance makes sense of the patterns of diversity/neutrality indexes and leads to an important observation that the divergence between eastern and western groups seems to have originated between central and southwestern Anatolia, during the Neolithic. In addition, the temporal tests detected no significant changes in haplogroup frequencies across Aşıklı Höyük’s occupation layers (P = 0.28 to 0.97).” ref
Variable kinship patterns in Neolithic Anatolia revealed by ancient genomes
“Genetic kinship estimated from co-buried individuals’ genomes in Neolithic Anatolia. Close relatives are common among co-burials in Aşıklı and Boncuklu. Many unrelated infants were found buried in the same building in Çatalhöyük and Barcın. Neolithic societies in Southwest Asia may have held diverse concepts of kinship. The social organization of the first fully sedentary societies that emerged during the Neolithic period in Southwest Asia remains enigmatic, mainly because material culture studies provide limited insight into this issue.” ref
“However, because Neolithic Anatolian communities often buried their dead beneath domestic buildings, household composition and social structure can be studied through these human remains. Here, we describe genetic relatedness among co-burials associated with domestic buildings in Neolithic Anatolia using 59 ancient genomes, including 22 new genomes from Aşıklı Höyük and Çatalhöyük. We infer pedigree relationships by simultaneously analyzing multiple types of information, including autosomal and X chromosome kinship coefficients, maternal markers, and radiocarbon dating.” ref
“In two early Neolithic villages dating to the 9th and 8th millennia BCE, Aşıklı Höyük and Boncuklu, we discover that siblings and parent-offspring pairings were frequent within domestic structures, which provides the first direct indication of close genetic relationships among co-burials. In contrast, in the 7th millennium BCE sites of Çatalhöyük and Barcın, where we study subadults interred within and around houses, we find close genetic relatives to be rare. Hence, genetic relatedness may not have played a major role in the choice of burial location at these latter two sites, at least for subadults. This supports the hypothesis that in Çatalhöyük and possibly in some other Neolithic communities, domestic structures may have served as burial location for social units incorporating biologically unrelated individuals. Our results underscore the diversity of kin structures in Neolithic communities during this important phase of sociocultural development.” ref
“This study focuses on social organization across two Neolithic periods. The Aceramic period is represented by Aşıklı Höyük (c. 8,350–7,300 cal BCE) and Boncuklu (c. 8,300–7,600 cal BCE) (Figure 1A), which are among the earliest sedentary communities in Central Anatolia. During the 9th millennium these sites were characterized by small curvilinear buildings, and both maintained mainly forager subsistence practices. The subsequent Ceramic Neolithic period communities were increasingly reliant on food production, and they lived in larger settlements characterized by rectilinear, clustered architecture. In our study, this later period is represented by Çatalhöyük (c. 7,100–5,950 cal BCE), Tepecik-Çiftlik (c. 7,500–5,800 cal BCE), and Barcın Höyük (c. 6,600–6,000 cal BCE). For this study, we screened Neolithic period human remains from Aşıklı Höyük (n = 30) and Çatalhöyük (n = 60) by shotgun DNA sequencing. Owing to adverse conditions and the antiquity of the material, only n = 8 (26%) and n = 14 (23%) skeletons (all petrous bones), respectively, contained ≥0.1% human DNA.” ref
Increased genetic diversity from the Aceramic to the Ceramic period
“We first analyzed genetic relationships at the population level. Principal components analysis (Figure 1B), ADMIXTURE analysis, as well as FST, f3– and D-statistics (Figures S2A) showed that Aşıklı Höyük and Çatalhöyük people belonged to the Central and West Anatolian early Holocene gene pool, represented by Boncuklu Höyük, Tepecik-Çiftlik, and Barcın Höyük individuals, as well as an Epipalaeolithic Central Anatolian individual from Pınarbaşı. Within this regional group, we discern genetically distinct communities, such that individuals from these sites (except for Tepecik-Çiftlik) tended to share more recent common ancestry with individuals from the same settlement compared with those of other settlements (among 576–11,780 D-tests per site, 84%–93% were nominally significant in this direction; Figures S3D). FST, f3– and D-statistics also showed that residents of the two Aceramic Neolithic settlements, Aşıklı Höyük and Boncuklu Höyük, were genetically highly similar to each other (Figures 1C, S2A, S2B, and S2D–S2F) relative to Ceramic Neolithic-period populations.” ref
“Aceramic Neolithic-period populations had lower within-group genetic diversity (measured using the f3-statistic) than did Ceramic Neolithic groups (Figures 1D and S2C, and Tables Z8 and Z9) and carried a higher fraction of short runs of homozygosity (ROH) than most Ceramic Neolithic genomes (Figure S3G). This temporal increase in diversity, also noted in earlier studies, could be explained by two non-exclusive scenarios, namely population growth and genetic admixture. By testing D(Outgroup, X; Aceramic Anatolian, Ceramic Anatolian), where X represents an early Holocene Zagros or Levantine population, we found results compatible with southern and eastern gene flow into Central and West Anatolia between roughly 7,500 and 6,500 cal BCE (Figure 1E and Table Z4) as previously suggested. Using qpAdm, we could also model Ceramic Neolithic Anatolian populations as mixtures of c.90% Aceramic Neolithic Anatolian ancestry (estimate ± 1 standard error: 89%–92% ± 2%–4%) and c.10% Levantine ancestry (8%–11% ± 2%–4%) (models that included Zagros or Caucasus populations were not supported) (Table Z10). Notably, the timing of increased population mobility is contemporaneous with a stronger reliance on agriculture and animal husbandry as food sources, a shift to larger buildings, likely population growth, and possible shifts in patterns of social organization, as we describe below.” ref
Estimating pedigree relationships among Neolithic co-burials
“Neolithic Southwest Asian settlements contain structures that are usually interpreted as domestic dwellings that served as focal points for the socialization of household members. These societies frequently interred their dead, including subadults and adults of both sexes, beneath the floors of these buildings while they were inhabited by the living. A common assumption has been that these burials were of household members who were related in some way, possibly genetically or through social kinship.However, it is not yet clear if individuals buried under house floors necessarily lived in those structures as part of a single co-resident group. The extent of dietary similarity among individuals interred within the same building, for instance, is currently ambiguous. Nevertheless, the assemblage of burials within or around domestic structures is expected to carry information about household composition and/or burial practices, and it may shed light on the relative importance of genetic relatedness as an organizing principle within these early Neolithic communities. In previous studies at Çatalhöyük, analyses of dental morphometrics and of mitochondrial DNA have suggested that individuals interred within the same building are often not genetically closely related. The question has remained unresolved, however, due to the inability of either data type to sufficiently identify exact pedigree relationships on any one site.” ref
“Here, we re-address the question of co-burial relationships using genome data from Neolithic Anatolian communities. In order to infer reliable pedigree relationships, we used different sources of information simultaneously. First, we employed three allele frequency-based methods to infer genetic kinship coefficients: NgsRelate, lcMLkin, and READ (Figures 2, S4A, and S4B). Second, to distinguish different pedigree relationships among putative first-degree pairs (e.g., siblings, mother-son, father-daughter), we used the probabilities of sharing 0, 1, or 2 alleles identical-by-descent (Cotterman coefficients; k0, k1, k2), although the low coverage of our genome data constrained the utility of this latter approach. Therefore, for inferring pedigree relationships we combined (a) kinship coefficients (θ) estimated from autosomal and from X chromosomal loci, (b) mitochondrial haplotype sharing, (c) osteological age-at-death estimates, and (d) radiocarbon dates. Finally, we performed pedigree simulations to determine the power of kinship coefficient estimation using low coverage data (Figure S4C). In addition, we studied the performance of the kinship estimation algorithms on negative controls, that is, real data from pairs of individuals who could historically not be close relatives. We hence limited the kinship tests to pairs of individuals sharing a minimum of 5,000 single nucleotide polymorphisms (SNPs) (Figure S1B). This permits reliable estimations of genetic relatedness up to the 3rd degree (e.g., cousins). Pairs related beyond the 3rd degree are here referred to as “unrelated.” ref
“The final dataset included a total of 223 pairs of individuals buried within the same sites, who were broadly contemporaneous, and who had sufficient genomic data for reliably inferring genetic relatedness (Tables S1, S3, and Z11). Of these, co-burials comprised 32 individuals and 50 pairs, including 2–6 burials associated with the same building or building clusters (i.e., co-burials). In Çatalhöyük and Barcın, co-buried individuals who could be genetically sampled only included subadults. Importantly, all these buildings either had evidence of domestic use (e.g., hearths) or lacked evidence of systematic non-domestic use (e.g., use as animal penning), and did not deviate from others of the same layer in terms of structure or elaboration.” ref
Co-buried pairs in Aceramic period sites frequently include relatives
“The data from Aşıklı Höyük included genomes of five individuals from the same stratigraphic layer who produced statistically consistent radiocarbon ages (χ2 = 7.6, χ 2(5%) = 9.5, n = 4; Table Z2) and could have lived at the same time. These individuals, all females, were interred in two buildings in close proximity and that shared a workspace, likely used by a single household (Figure 3A). All three methods identified two pairs of first-degree relatives (Figure 2A, and Tables S3 and Z11). One pair buried in the same building included an adult and child (individuals 136 and 131). The other pair, buried in separate but proximate buildings, included an old adult and child (individual 133 and 128). The genetic and skeletal evidence indicated both pairs to be sisters (Figures 2A–2C and Tables S3 and Z11). However, we cannot exclude parent-offspring relationships. An adult female (individual 129), buried in the same building as individual 128, had no genetically close relatives among the other four individuals. Thus, although only a minority of studied individual pairs (2 of 10 pairs) were closely related, the majority of individuals studied (4 out of 5) had one close relative identified in the same or adjacent building (Figure 2D and Table S1).” ref
“The Boncuklu Höyük data comprised nine genomes of individuals who were buried in three buildings or in external spaces. Five individuals formed a co-burial cluster in two adjacent consecutive buildings (Figure 3B). Among these, two pairs of first-degree relatives were identified (Figures 2A–2C; Tables S3 and Z11) (also reported earlier). The first was a possible mother and her adult son (individuals ZHF and ZHJ), who were buried in the same building (B14). Their radiocarbon results were different at the 1% significance level (χ2 = 8.8, χ2 (5%) = 3.8, n = 1; Table Z2), and suggested that the woman (ZHJ) died first with 90% probability. The second included a possible pair of adult male and female siblings (individuals ZHBJ and ZHAF). These individuals were buried in the proximate consecutive buildings (B12 and B14). Thus, as at Aşıklı Höyük, we could identify close relatives across the majority of individuals (4 out of 5) associated with neighboring building pairs (Figure 2D and Table S1). The only exception was a perinatal infant (individual ZHAG). Intriguingly, this infant was buried in the same grave with an adult female (individual ZHAF). The infant also shared the woman’s mitochondrial haplotype but was closely related to neither the woman nor any other individual studied. Other individuals also lacked close relatives in this dataset.” ref
Relatives are rare among Çatalhöyük and Barcın intramural burials
“The Çatalhöyük data contained genomes of 14 individuals from multiple stratigraphic levels. All except one individual were subadults; 10 and 4 were genetically determined to have been females and males, respectively. Ten subadults, buried in three buildings dating to the mid-7th millennium BCE, constituted three co-burial clusters (Figure 3C). We identified a single pair of female siblings (individuals 2728 and 2842), an infant and a child, buried within the same building (Building 50) (Figures 2A–2C, and Tables S3, Z2, and Z11). The pair produced statistically consistent radiocarbon measurements (χ2 = 0.0, χ2 (5%) = 3.8, n = 1). None of the other pairs of individuals tested were closely related. Hence, among Çatalhöyük individuals co-buried in these three buildings and tested genetically, only 2 out of 10 had genetic kin identified (Figure 2D and Table S1).” ref
“The Barcın Höyük data included genomes of 23 individuals from multiple phases (VIa, VIb and VIc or VId2/3) (Figure 3D). Ten of these individuals were inserted into three or possibly four buildings (Table Z2). We determined two pairs of relatives, including a pair of subadult sisters (associated with Building 5) and a pair of subadult males who were second- or third-degree relatives (associated with Buildings 14/15) (Figures 2A–2C and Tables S3 and Z11). Both pairs were buried in close proximity to each other and produced statistically consistent radiocarbon measurements (L11 213 & 215, χ2 = 0.7; M10 271 & 275, χ2 = 0.2, χ2 (5%) = 3.8, n = 1 for both; Table Z2). None of the other individuals had close relatives identified, including four infants buried in Building 4. Hence, among co-buried individuals we could identify relatives for only 4 out of 10 (Figure 2D and Table S1).” ref
“The Tepecik-Çiftlik data included genomes of a total of five individuals from two strata. We identified a probable pair of a mother and her adult son (individuals 37 and 21) buried in different parts of the same building (Building AY/AK) (Tables S3 and Z11). These individuals produced radiocarbon results that are different at the 1% significance level (χ2 = 8.0, χ2 (5%) = 3.8, n = 1), which suggests that it is 96% probable that the woman (individual 37) died first (Table Z2).” ref
Temporal or age-dependent variability in co-burial kinship patterns
“The identification of multiple instances of close genetic relatedness among co-burials across all Neolithic Anatolian settlements studied suggests that early Neolithic social arrangements and possibly household composition were to some extent linked to genetic ties. Although long assumed, genetic relatedness within Neolithic house-related social groups is documented here directly for the first time. This is particularly salient in the evidence from 9th and early 8th-millennium BCE Aşıklı Höyük and Boncuklu Höyük and could be considered suggestive of elements of close genetic kin relationships among groups buried together within Aceramic Neolithic houses.” ref
“Nevertheless, a notable fraction of our sample also contained individuals (nearly all subadults) buried in buildings together with genetically unrelated individuals (50% of 32 individuals; Figure 3). Genetic relatedness among co-burials was especially low in the 7th-millennium BCE Çatalhöyük and Barcın Höyük, with the majority of co-burials lacking identifiable genetically related kin (the sample size from Tepecik-Çiftlik is too small to reach a general conclusion). Indeed, the combined frequencies of individuals among co-burials with and without identified relatives appeared different between Aşıklı and Boncuklu versus Çatalhöyük and Barcın Höyük (odds ratio = 8.6, Fisher’s exact test p = 0.019; Figure 2D and Table S1). However, the difference becomes non-significant when including the co-buried adult pair from Tepecik-Çiftlik in the temporal comparison between Aceramic and Ceramic period sites (odds ratio = 6.6 and p = 0.054).” ref
“Two points need further mention. First, although all age groups are represented archaeologically among Çatalhöyük and Barcın Höyük burials, among samples with sufficient DNA data we had high proportions of subadults (13/14 and 16/23, respectively). This effect appears to be caused by better DNA preservation in subadult bones, at least at Çatalhöyük (Figure S1C; STAR Methods), possibly as a result of age-based differences in burial treatment. As a consequence, in our study, no adult co-burials could be genetically examined from these two sites. Second, Çatalhöyük and Barcın Höyük buildings were significantly larger and contained more burials than those of the Aceramic Neolithic sites (Figure 3).” ref
“The infrequency of close relatives among subadults buried together in relatively large structures at Çatalhöyük and Barcın Höyük is intriguing. It raises the question of whether these buildings may have been used by extended families, such that the co-buried subadults could be distant cousins who were not identified by the methods employed. We thus tested whether individuals buried in closer proximity shared greater genetic similarity, using genetic distances based on the f3-statistic (STAR Methods). After excluding close relatives, we found no correlation between genetic distance and spatial distance across burial pairs in either Çatalhöyük or Barcın Höyük (Pearson r < 0.02, Mantel test p > 0.3; Table S4).” ref
“Researchers also tested the hypothesis that overall genetic similarity among co-burials might be higher within buildings than between buildings. Again, we found no evidence for this (one-sided permutation test p > 0.8; Table S4). These results corroborate previous analyses that found no significant correlation between burial location and dental similarities in Neolithic Çatalhöyük adults and also a lack of mitochondrial DNA shared among co-burials. We note that we do not expect all individuals associated with these buildings to have been buried within those structures. Also, not all individuals interred in these buildings could be sampled in this study. Still, the presence of individuals without identified relatives implies that the choice of the same structure for the burial of community members may be motivated, among other factors, by additional forms of social connectedness.” ref
“For instance, co-burials, including juveniles, may have included “adoptive, foster or fictive kin held together by memory and history-making.” Accordingly, co-burial and perhaps household composition in these later Neolithic settlements may have included—but also extended beyond—close genetic kin. It is also possible that the practice of co-burying subadults with genetically unrelated individuals was already present in the Aceramic period in Anatolia, but we did not sample these sufficiently in Aşıklı and Boncuklu. Indeed, the Boncuklu adult female-infant pair sharing a grave, found to be unrelated, may reflect such a tradition. It, therefore, remains unclear, yet, whether the difference among sites in co-burial patterns reflects a temporal shift or differential treatment of adults versus subadults in Neolithic Anatolia.” ref
Varying traditions linking sex and space
“Another set of observations involves burial patterns with respect to sex. First, we find co-burial of closely related adults of both sexes at Boncuklu Höyük and possible adult-child sister pairs at Aşıklı Höyük. Although our sample size is limited to reach a definitive conclusion, it is worth noting that the pattern is consistent with adult females retaining close ties to their natal households, symbolically or residentially, over significant periods of their lives. This scenario, at least at Boncuklu Höyük, could equally have applied to the males. Second, the sex patterning observed in Anatolian Neolithic burials appear distinct from those described for Neolithic and Bronze Age cemeteries in Europe, where male burials predominate, and patrilocality is evident.” ref
“For instance, in a study of multiple cemeteries, Mittnik and colleagues identified only 2 first-degree related female pairs out of 21 first-degree relationships. This proportion is different in our data, which reveals 4 first-degree related female pairs out of 7 first-degree relationships (odds ratio = 11.1, Fisher’s exact test p = 0.02). This result, as well as the contrast between co-burial of related adult females in the Aceramic period buildings and the stark patrilocal patterns in 6th-3rd millennium European cemeteries, are consistent with the notion that sex role differences intensified following the initial adoption of agriculture. Meanwhile, both sister pairs we identified at Barcın and Çatalhöyük were subadults. In this regard, patrilocal traditions in Ceramic period Anatolian sites remain a possibility (as suggested earlier based on dental and mtDNA data).” ref
“In summary, in addition to evidence for the existence of close genetic ties among putative households in the Aceramic period, we find that genetic relatedness among subadult co-burials was infrequent at Ceramic period Çatalhöyük and Barcın. Although we cannot yet pinpoint when and where this latter practice emerged, it appears plausible that during the transition from the Aceramic to the Ceramic Neolithic period in Anatolia, in parallel with changes in subsistence and population mobility, genetic relatedness may have become less important in the structuring of intramural burial traditions.” ref
Neolithic buildings and households
“The concept of “house” refers to a social institution through which societies define a particular type of membership group, i.e., the “household.” What defines a household is based upon the cooperating individuals’ criteria for relatedness, task-orientation, and co-residence. These criteria are socio-culturally constructed and, therefore, highly variable across societies. For example, household members can be genetically related, as in genetic kin-based family organizations, but a household can also be composed of individuals who co-reside and share tasks with reference to relatedness criteria other than genetic ties. Nevertheless, these criteria of relatedness, genetic or otherwise, are considered legitimate only if they express continuity through successful invention and manipulation of concepts such as descent, belonging and other social differences based on age, sex and skill, all of which are also actively employed in terminologies of kinship or affinity. Within this context, long-lasting architecture has been the most potent embodiment of inclusion and relatedness, through which a household membership and its history can be represented via a variety of symbolic activities.” ref
“Some of the earliest long-lasting residential architecture, considered to be the primary context for the socialization of household members, is found in early Neolithic SW Asia, c. 10th-7th millennia cal BCE. The criteria that define relatedness among the household members of these societies, however, have long been debated: were the co-residents genetic kin, or did other factors determine household membership? Based on the size and form of the buildings, it has been suggested that the earlier curvilinear structures of the c.10th-9th millennium cal BCE were used by extended families, perhaps related to polygamous social structures, whereas the adoption of larger rectilinear and compartmentalized buildings of 8th-7th millennium cal BCE reflects a shift to close genetic kin-based organization.” ref
“Alternatively, given the relatively small size of most Neolithic residential structures, regardless of shape, it has been postulated that these buildings were mostly used by nuclear families. Other researchers hold that the transition from some form of nuclear family household to increasingly autonomous family households occurred during the Pre-Pottery Neolithic B (PPNB). Yet others argue that the increasingly autonomous households only occurred in the Late Neolithic as an element of multiscalar transformations of Neolithic communities in this period. Research on mortuary practices also underlines the broad regional changes through time, including suggested shifts from community membership to increasingly separate and autonomous household organizations in the PPNB. Meanwhile, the repeated construction of mudbrick buildings at the same location over multiple generations, sometimes even maintaining the position of internal structures such as hearths, implies the presence of distinct household identities in these societies.” ref
Do co-burials represent households?
“One potential source of information that could help resolve the nature of Neolithic household composition and social organization comes from burials within buildings during their occupancy. Neolithic SW Asian societies frequently practiced the burial of individuals beneath the floors of domestic buildings, usually during the time these structures were inhabited. A common assumption has been that these burials were of household members and were related in some way, possibly genetically or through kinship based on other factors. This could include households composed of families of closely genetically related individuals, extended families, multi-family households, or social units where genetic relatedness had little role. In reality, however, it remains unclear whether individuals buried under house floors lived in the same building as part of a co-resident group, i.e., whether they represented households.” ref
“If co-burials were indeed household members, we may expect them to share specific attributes more with each other than with other co-burial groups; most notably, elements of their diet. Evidence on dietary similarity among Neolithic Anatolian households is currently equivocal. A 2015 study reported no dietary differentiation among Çatalhöyük co-burials in different buildings. A 2020 study using a wider dataset again from Çatalhöyük reported statistically significant differentiation in carbon and nitrogen isotope values among buildings. This same work further reported significant dietary differences among neonates buried in different buildings. Still, possible confounding factors that could influence stable isotope profiles (age and sex for adults, pathological conditions for neonates) were not explicitly controlled for in these analyses, and we therefore consider these results as preliminary.” ref
“There exist additional arguments against the hypothesis that co-burials represent households. It appears that the average number of burials per residential structure is generally too small to represent full households. For instance, in Aşıklı Höyük, only 90 burials have been discovered from more than 400 rooms excavated. This suggests additional factors influenced the choice of burial locations and type of funerary treatments of individuals. Furthermore, an apparent excess of burials in some residential buildings, in sites such as Çatalhöyük, and occasionally at other sites such as Abu Hureyra and Bestansur (although the relevant buildings here may not be ordinary residential structures), implies a special role of some residential buildings for burial of individuals who probably had originally lived in other residences. Düring and Marciniak’s (2005) analysis of Çatalhöyük houses also indicates that human burials in buildings may have served to advertise the temporal continuity (history) of the buildings, which thus ensured the continuity and success of the household, regardless of their genetic ties.” ref
“If co-burials do not represent household members, their interment in the same buildings could be driven by at least two distinct traditions. First, individuals may be buried together because they died at the same time. This could also include mass burials following disease outbreaks. However, in the case of co-burials in Neolithic Anatolian settlements, the mortuary context and mortality profiles do not indicate mass burials. The evidence overall suggests these were collective burials, where individuals were buried sequentially, as is prevalent at Neolithic Çatalhöyük as well as other sites.” ref
“Second, co-burial patterns may reflect local traditions stipulating specific burial arrangements of individuals who do not belong to the same household. Such traditions could involve burying individuals of specific status or social backgrounds together. The motivation behind these traditions may be to maintain social and economic ties among groups and to “consolidate community membership”. For instance, it has been suggested that the emergence of cemeteries during the Natufian period could have represented “the establishment or strengthening of special interest groups, inheritance of corporate property, and territorial ownerships”. Another example could be traditions such as described for Aboriginal Australian groups where the corpses of deceased young children were retained by the mothers to be interred with an adult male who dies next (Musgrave 1930, cited in). If such arrangements were in place also in Neolithic Anatolian settlements, we might expect no direct social or genetic connection among co-burials.” ref
Relatedness among co-burials
“Studies on genetic relatedness among co-burials in Neolithic SW Asia have yet been limited. Most work to date relies on dental metric and non-metric traits as proxies for genetics, and one recent study used mitochondrial DNA. These studies have reported patterns consistent with endogamy or with matrilocality in Neolithic Levantine sites, and with patrilocality at Çatalhöyük. Meanwhile, the Çatalhöyük studies, based either on dental analysis or mitochondrial DNA, found no evidence for individuals buried in the same building being more closely related to each other than to individuals buried in other buildings.” ref
“Still, owing to the inability to estimate the degree of kinship using dental traits and mitochondrial data, the question of kinship among co-burials in Neolithic SW Asia has remained largely unresolved. Ancient genomics, in turn, can be used as a powerful tool to determine genetic relatedness and kinship among households of the dead, allowing further consideration of how burial locations might have structured relationships between households of the living and the construction of kinship, as well as social memory and social traditions in general. With some temporal depth to our study we are also able to consider if there might be temporal trends in these social practices over the long term.” ref
Description of archaeological sites
Description of Aşıklı Höyük
“Aşıklı Höyük, located in the Volcanic Cappadocia Region in eastern Central Anatolia is one of the earliest sedentary communities of the region, radiocarbon dated to the mid-9th and 8th millennium BCE (8350-7300 cal BCE). Excavations at the site started in 1989 as salvage excavations under the direction of Prof. Ufuk Esin (İstanbul University). Since 2010, the research and excavation project has been led by Prof. Mihriban Özbaşaran (İstanbul University) and Güneş Duru (Mimar Sinan Fine Arts University) in collaboration with an international team from various universities and institutions.” ref
“The first inhabitants of Aşıklı settled near the western bank of the Melendiz River. The river, flowing from the Ihlara Valley, and the volcanic landscape provided a rich habitat for various animal and plant species. A warm climate and park-woodland vegetation was dominant in the region during the beginning of the Holocene. The mid-9th millennium BCE inhabitants of the site lived in semi-subterranean, oval mudbrick buildings that were reconstructed and renewed periodically at the same location. Characteristics of these buildings include hearths, a small platform, grinding stones and burial pits. Daily life was organized outside the buildings, in open activity areas, where many of the daily tasks were conducted.” ref
“Archaeozoological data attest to broad spectrum hunting during the 9th millennium BCE, including a variety of small prey animals, birds and fish, although the main focus was always on sheep/goat. Analyses of micromorphology and soil chemistry, and the presence of primary dung layers attest to the fact that animals were kept on-site, inside wattle and daub enclosures. Archaeozoological data, as well as isotope analysis show that caprines, specifically sheep, were kept in the settlement from the earliest levels; management and the domestication process continued all through the sequence. The community had the knowledge and the experience of growing plants and cultivating wild and domestic cereals. Wild plants, legumes and fruits were among the gathered plants.” ref
“With the start of the 8th millennium BCE, changes took place in architecture and settlement patterns. Rectangular structures replaced the oval and semi-subterranean buildings. These rectangular buildings were mostly single-roomed. Although few in number, buildings with two or three-room buildings are also present. Toward the end of the settlement occupation, buildings started to cluster. Building clusters, generating neighborhoods, were separated by narrow spaces or passages with access to communal middens. Separated by a “gravel street” from the residential area, to the southwest of the present mound, lies a building complex distinguished from domestic buildings in terms of its plan, size, construction material, internal architectural features and floor and wall treatment. The architectural features and the characteristics of the archaeological material (i.e., the dominance of wild cattle) permit interpretation of this area as a “public area” where communal consumption and certain ceremonies took place. Evidence of communal activities in this area indicates the continuity of the collective way of living, while the daily activities in the residential area most probably took place on the flat roofs and inside the dwellings. During this period, hunting and gathering continued, though with less importance, and subsistence was based mainly on sheep and goat, but these animals were no longer kept within the settlement.” ref
“Two concepts, a communal way of life and continuity, characterized the social organization of the Aşıklı community. Interaction with other regions and communities had a certain tempo during the mid-9th millennium BCE, as evidenced by the material culture. However, simultaneous with the increasing focus on the full establishment of sedentism and caprine management, the pace of interaction decreased, only to increase again during the last 200-300 years of site occupation, corresponding roughly to the second half of the 8th millennium BCE. This is well illustrated by the sudden appearance of non-local materials and technologies during this period. In contrast with this pattern of temporal change, continuity of certain elements, such as the location of the buildings and interior architectural features, constant renewal and maintenance of the floors and walls of buildings, and the transferring of objects and know-how was another factor that characterized the social fabric of the community. The inhabitants managed to live in cohesion throughout the occupation sequence and the communal way of life was maintained with new solutions, but also continuity through temporal changes and transformations was the main characteristic of the newly established Neolithic way of life at Aşıklı Höyük.” ref
“The burial customs consist of intramural, single sub-floor inhumations. The deceased were buried in pits under the floors of the buildings in a flexed position. To date, 90 burials have been found in 400 rooms. Although this tradition was not subject to change for hundreds of years, new practices arose during the latest levels of the occupation at the site. The dead were not buried with any items of personal adornment during the mid-9th millennium BCE. However, changes can be observed toward the mid-8th millennium when some individuals are found buried with ornaments. Of the 82 individuals subjected to bioarchaeological analysis, adults constitute 60% while children make up 40%. Of the 46 adults for whom sex can be determined, females constitute 65% while males constitute 35%, a marginally significant difference (binomial test p = 0.054). In terms of the daily activities conducted by the Aşıklı Höyük individuals, task-related pathologies of adults show that the shoulders, hips, ankles, elbows and knees were affected by osteoarthritis, possibly stemming from habitual stress. Males exhibit significant degrees of shoulder osteoarthritis, followed by their elbows and hips; for females the ankles were most affected by this disease, followed by the shoulders and hips. This may suggest that males were routinely engaged in activities such as carrying heavy loads, throwing, walking and kneeling, and females were probably engaged in activities that involved walking and squatting.” ref
“Five of these burials genetically studied here were interred in Building 1 and Building 3 of Aşıklı Höyük Layer 4 (Figure 3A). These are buildings in direct proximity with less than 1 m between them, which showed temporal overlap in their periods of use, and which shared a common open workspace between them. We therefore treated the individuals from both buildings as a cluster of co-burials, who might represent members of the same household.” ref
Description of Boncuklu Höyük
“Boncuklu is situated in the middle of the SW Konya basin (37°45’N 32°52’E) and lies 33.4 km northwest of the site of Pınarbaşı and 9.5 km northeast from Çatalhöyük. The site was discovered during the archaeological survey under the direction of Prof. Douglas Baird from the University of Liverpool, UK. Excavations directed by Baird began in 2006 and continue at the present time. Baird was joined by co-directors, Prof. Andrew Fairbairn of University of Queensland, Australia and Dr. Gökhan Mustafaoğlu of Ankara Haci Bayram Veli University, Turkey, since 2011. Occupation of the site is documented from 8300-7600 cal BCE directly through radiocarbon dating. However, stratigraphic and material evidence suggest a slightly longer span of occupation.” ref
“The exploitation of wild resources seems to have predominated, especially wild cattle and boar, fish and wetland birds, along with nuts and fruits from surrounding hill areas. Small-scale cultivation of wheat, lentils and peas was an additional modest component of subsistence activities. The chipped stone industry was microlithic, in significant contrast to broadly contemporary Levantine PPNB and northern Fertile Crescent assemblages and thus shows significant continuities with the earlier, local Epipalaeolithic and the earlier 10th/early 9th millennium BC community at Pınarbaşı in technology and raw material. Continuities between Epipalaeolithic and early Holocene forager communities and the community at Boncuklu are clear. This evidence is supported by significant genetic continuity. By 8300 cal BCE it appears local foragers adopted domestic plants from areas to the south and east, incorporating them into their traditional wetland exploitation practices. They were probably introduced to the region as a consequence of the far-reaching and continuous interactions with neighboring regions from the Epipalaeolithic through the 10th-early 9th millennia cal BCE, as also documented at earlier and contemporary Pınarbaşı.” ref
“The site possessed a number of sunken-floored sub-oval domestic buildings with mudbrick walls. The households display highly structured use of internal house space, divided into a ‘clean’ area presumably for sleeping, socializing and food consumption and a ‘dirty’ kitchen area. The houses were very regularly refurbished, plastered and modified, especially the hearth areas, showing the intensity of domestic use. The floor area of these houses is small and modeling shows small intimate household units with intensive and repetitive domestic practices. Evidence of ritual and symbolism in the ‘clean’ areas, including burials, is regular and differentiated from house to house suggesting creation and maintenance of distinctive household identities. The Boncuklu houses were also repeatedly and continuously reconstructed over multiple generations in the same location, a practice at some other 10th-7th millennia cal BCE sites in the surrounding regions, for example to the northeast at Aşıklı from 8300 cal BCE, just to the south at Çatalhöyük from 7100 cal BCE, in the Levant at PPNA Jericho and in PPNB Tell Halula. This seems to be a symbolic statement of household continuity. This expression of continuity and identity suggests small tight-knit households in continuous occupation of these domestic structures, whatever the nature and dynamism of their composition, which we can start to grasp through aDNA evidence. Nevertheless, there seems evidence that some broader corporate social practices cross-cut households, including some practices involved in food and resource exploitation in the wider landscape.” ref
“Primary inhumations were placed under the ‘clean’ area of the houses during their occupation. It seems the dead ‘ancestors’, whether biologically related or not, were kept close to the living. In the case of Boncuklu the modest numbers of burials under house floors, maximum 5 and more usually 1-3, per house, suggest many of these could easily be members of the household that lived in these buildings, although we certainly cannot assume that to be so. Nevertheless, reflecting the fact they occurred within the houses while still in use and that these were small-sized buildings with very intimate spaces, presumably means the co-burial of the dead expressed some type of relationship to the households of the living, and thus represented a symbolic statement of connection between the dead and the living. Indeed, evidence attests to ongoing attention to burials and knowledge of their location.” ref
“There were also primary burials and burials of deliberately disarticulated human remains, including human crania, in open areas between buildings in areas of midden accumulation. More than 37 Neolithic burials, plus a minimum of 274 individual bones and 129 isolated finds of human remains have been studied, although more have been excavated. Nine skeletal samples from securely stratified 9th-8th millennia cal BCE burials in Areas H, K, and M provided sufficient aDNA preservation for genetic analysis (Table Z2), and thus genomic data. Boncuklu human remains do not reflect significant disproportionate representation of males or females and there is an even spread across most age categories, including children and young, middle and old adults, with a slight, but not unusual, lesser presence of older children/adolescents.” ref
“Five of these burials (ZHF – Grave 14, ZHJ – Grave 15, ZHAF – Grave 18, ZHAG – Grave 18 and ZHBJ – Grave 30), including 2 pairs of individuals with first-degree genetic relationships were all articulated primary inhumations stratified within a sequence of 2 buildings in Area H, Building 12 and Building 14. Building 12 predates Building 14 and, indeed, the foundation cut for Building 14 removed the northern edge of Building 12 (Figure 3B). Building 14 seemed a direct replacement for Building 12, an example of the continuous reconstruction of the buildings in the same locations, although in this case with some shift of the house to the north. ZHBJ, the likely brother of ZHAF, was buried in the northern part of Building 12 (Figure 3B). ZHAF, his likely sister, was buried in the southern part of Building 14 (Figure 3B). This seems a deliberate attempt to keep these individuals close at death and points to the close connections between the living and dead in these households. Both these burials had similar orientations, approximately west-east/northwest-southeast with heads at the West. It is thus tempting to think this might also reflect their close relationship.” ref
“It may well have done, but these are the most common burial orientations at Boncuklu, among c. 70% of the analyzed burials and so might simply reflect these broader patterns. ZHAG, a female perinatal child that likely died at birth, was placed directly against the pelvis of ZHAF, but was genetically unrelated to that adult female ZHAF and also unrelated to ZHF and ZHJ in the same building. It is, of course, possible her mother lived in Building 14 but was genetically unrelated to the other adults buried there, or that as a result of some form of connection to the child and/or her mother she was buried with ZHAF, albeit from a household who lived elsewhere. ZHF and ZHJ, most likely adult son and mother respectively, were located in the more eastern parts of Building 14 (Figure 3B). The orientation of their bodies was not dissimilar, ZHF had the common northwest-southeast and ZHJ a north-south orientation. However, their heads were at opposite ends of the grave-cut, ZHF to the northwest and ZHJ to the south. It is, therefore, difficult to suggest that orientation at Boncuklu was a direct expression of close family relationships.” ref
“ZHAJ was a primary inhumation burial of an adult male that predated Building 14 and seems to have been located in an open area. ZHB was the burial of an adult female burial post-dating Building 14. Overlying stratigraphy was eroded so it was unclear whether the grave for ZHB was cut through the floor of a building or was placed in an external area. These two burials do not show any close genetic relationships to the other sampled individuals. The other burials analyzed, genetically unrelated to any of these burials in Area H, was one adult male primary inhumation, ZKO, buried in Building 9 in Area K, broadly contemporary but c. 15 m from the cluster in Area H. ZMOJ was a primary inhumation in an external area in the middens of Area M, located c. 25 m from the cluster in Area H. Although well stratified in Neolithic deposits the chronological relationship with the Area H cluster and ZMOJ is not clear.” ref
Description of Çatalhöyük
“Located 9 km to the south of Boncuklu Höyük on the Konya Plain in Central Anatolia, the site of Çatalhöyük was discovered and first excavated between 1961-1965 by James Mellaart (British Institute of Archaeology at Ankara), and later between 1995-2015 by Ian Hodder (Stanford University). Çatalhöyük was designated a UNESCO World Heritage Site in 2012. The site consists of two separate mounds or “tells.” The larger East Mound covers an area of 13 ha and has been dated to c. 7100-5950 cal BCE, corresponding roughly to the Ceramic Neolithic period. The smaller West Mound dates to the Early Chalcolithic and was occupied until the middle of the 6th millennium BCE. The Neolithic East Mound, until c. 6300 cal BCE, is characterized by dense clusters of mudbrick domestic structures interspersed with external spaces used for refuse disposal, animal penning and other daily activities.” ref
“To date, large-scale, clearly identifiable public structures have not been documented at the site. Instead, individual houses at Çatalhöyük appear to have served as the focal point not only for domestic activities such as craft production, food storage and processing, but also ritual behaviors such as burials, wall paintings and other architectural embellishments associated with an elaborate symbolic repertoire and reflecting a complex socio-cultural environment. There is ample evidence for the cultivation of domesticated cereal crops and the keeping of domesticated sheep and goats at the site. Wild animal species, including aurochs, also formed part of the diet, and in the later occupation phases (6500-5950 cal BCE) there is evidence for the herding of domesticated cattle.” ref
“Between 1993 and 2017 the skeletal remains of over 700 individuals had been recovered from stratified Neolithic contexts at Çatalhöyük. Primary inhumations (n = 471 individuals) placed beneath the floors of houses are the dominant burial type at the site. Individuals were typically buried in narrow oval pits under the eastern and northern platforms of the central room, although prenates, neonates and infants were also recovered from within side rooms and near ovens and hearths. Secondary burials of loose or partially articulated skeletal remains, often in association with primary burials, are also observed, although less frequently. Intramural burials became increasingly rare toward the end of the occupation of Çatalhöyük East, while burials are almost completely absent within the settlement on the Chalcolithic West Mound.” ref
“Of the 471 individuals from primary burial contexts, there are 178 adults (20+ years), 29 adolescents (12-20 years), 90 children (3-12 years), 67 infants (2 months-3 years), 85 neonates (0-2 months), and 22 prenates (> 38 weeks in utero). Among the adults and adolescents whose sex could be determined (n = 155), 89 individuals (57%) were assessed as females or possible females, while 66 individuals (43%) were assessed as males or possible males, a marginally significant difference (binomial test p = 0.077).” ref
Description of Barcın Höyük
“Located in the Yenişehir Valley in the province of Bursa in northwestern Turkey, the site of Barcın Höyük yielded an uninterrupted stratigraphic sequence from 6600 to 6000 cal BCE. The settlement was built on a low natural elevation in what would have been a marshland valley. The Neolithic levels at Barcın Höyük, which lie beneath a relatively thin deposit of later levels dating to the Chalcolithic, Bronze Ages and the Byzantine period, are thick and exceed 4.5 m in most places at the site. Called level VI, the Neolithic phase is divided into seven subphases: VIe (earliest level) through VIa. The VIe levels of the site represent the earliest farming community known to date in the Marmara Region. The initial pioneer communities who arrived here around 6600 cal BCE brought with them crops to cultivate and animals to herd. With regards to plants, domesticated varieties of cereals and pulses were plentiful. Sheep and cattle were the preferred herd animals although goats were also present while hunting only contributed a minor part of the diet. Extensive organic residue analyses on pottery demonstrate that the inhabitants of Barcın Höyük relied heavily on dairy products. This observation confirms those made by Evershed and colleagues for later sites in the Marmara Region.” ref
“The initial settlers in the region were accomplished potters even though pottery use was initially limited and indirect methods of heating foods were preferred. Within a century however, thin-walled finely made burnished pots become plentiful. Building on a consistent tradition, recipes of manufacture and temper changed over the ensuing centuries. The residents of Barcın Höyük lived in rectilinear timber frame houses with wood and mud walls. Houses tended to be in rows, surrounded by courtyard areas where a variety of activities were carried out. Burials associated with the settlement were placed within and near structures. Interestingly, many infants were buried within the house proper beneath floors while adults were typically placed in the courtyard areas. Children often tended to be buried outside but closer to the structures, sometimes beneath the floors of the verandas in front of the houses.” ref
“Although intensively analyzed for DNA, the Barcın Höyük skeletons await final anthropological analyses. Based on preliminary data, adults appear to comprise 38% (46 burials) of the 121 burials that come from primary burial contexts. Of the skeletons that can be identified based on sexual characteristics, nearly two thirds of these appear to be females or possibly females. Subadults including adolescents, children, infants and neonates comprise the remaining 62% of the assemblage.” ref
Description of Tepecik-Çiftlik
“Tepecik-Çiftlik is located in the Volcanic Cappadocia region of Central Anatolia in the Melendiz/Çiftlik Plain. The excavators suggest it was occupied from the end of the Aceramic Neolithic Period until the early Chalcolithic Period, between c.7500-5800 cal BCE. The Pottery Neolithic levels show evidence of agriculture and animal breeding, as well as continued hunting and gathering. The site is in close proximity to major obsidian ore beds in the region and is notable for its large amount of obsidian tool remains. Further information about the site may be found at. A 2016 report on Tepecik-Çiftlik indicated that over 170 individuals’ remains dating to the Neolithic levels, buried inside buildings and in open areas had been excavated. A collective burial was also found, and is thought to have been used for successive burials, both primary and secondary. It includes at least 42 individuals of both sexes and various ages.” ref
Description of archaeological material
“This section describes bioarchaeological characteristics of the individuals from Aşıklı Höyük, Çatalhöyük and Boncuklu Höyük. Some of this data are unpublished. Barcın Höyük and Tepecik-Çiftlik individuals included in this study have been described in the supplementary material of Mathieson et al. and Kılınç et al., respectively. Sex was estimated using dimorphic markers, and individual ages-at-death were estimated using standard methods such as human growth and epiphyseal fusion, dental calcification, and bone maturity/size. The sex of subadult individuals listed below have been determined based on genetic data produced in this study.” ref
Description of Aşıklı Höyük individuals
“SK2 (Level 1/2A; Building AB): the burial of a young adult female. Double burial. SK2 was buried in the same burial pit of a male, slightly later. The pit is located in a one-room rectangular building of the mid-8th millennium BCE settlement. Radiocarbon dating places the individual to 7585-7475 cal BCE (Table Z2). SK33 (Level 2C, Building C): the burial of a child, buried under the floor of a rectangular planned kerpiç (mudbrick) building. Radiocarbon dating places the individual to 7945–7890 cal BCE (9%) or 7870–7595 cal BCE (86%). Building C was renewed 10 times at the same location (Figure 3A), where this child’s burial was contemporary with its eighth renewal phase. Excavated in 1991. SK40 (Level 2B, Building BH): the burial of an old adult female. Sub-floor inhumation in a rectangular kerpiç building of the 8th millennium BCE settlement. One of the three individuals buried in the same building: a one-month old infant and a middle adult female. Radiocarbon dating places the individual at 7935–7915 cal BCE (1%) or 7825–7590 cal BCE (94%).” ref
“SK128 (Level 4, Building 3): the burial of a female child. She is one of the two individuals buried in the same building. Radiocarbon dated to 8225–7955 cal BCE (95%). SK129 (Level 4, Building 3): the burial of a young adult female, buried in a semi-subterranean oval building. She is one of the two individuals buried in the same building. Excavated in 2011; primary burial; radiocarbon dated to 8170–8115 cal BCE (6%), 8060–8045 cal BCE (1%), 8010–7985 cal BCE (1%), 7970–7735 cal BCE (86%). SK131 (Level 3E/4, Building 1): the burial of a female child, exposed lying on the pavement of a hearth in a semi-subterranean oval building. This is an exceptional burial, in position and in location. Four more individuals were buried in the same building. The burial was exposed in 2012. She was radiocarbon dated to 8200–8110 cal BCE (16%) or 8095–8035 cal BCE (7%) or 8015–7740 cal BCE (72%).
“SK133 (Level 3E/4, Building 1): the burial of an old adult female, the oldest member of the community thus far excavated. She was one of the five individuals buried in the same oval, semi-subterranean building, B.1. She was a primary burial and was radiocarbon dated to 8170–8115 cal BCE (8%), 8060–8040 cal BCE (1%), 8010–7980 cal BCE (2%), 7975–7735 cal BCE (84%). Excavated in 2012. SK136 (Level 3E/4, Building 1): the burial of a young adult female, one of the five individuals from Building 1. She was a primary burial, and was radiocarbon dated to 8175–8110 cal BCE (7%) or 8090–8075 cal BCE (1%) or 8065–8040 cal BCE (1%) or 8015–7705 cal BCE (84%) or 7695–7655 cal BCE (2%). Excavated in 2015.” ref
Description of Çatalhöyük individuals
“Sk.5357 (burial feature 576, Level South K, Early period, Building 17): primary burial of a male infant. He was 9 months ± 3 months at death based on dental development. It was buried in a flexed position along the east wall of B.17 in association with red pigment and traces of reed basketry. The burial was excavated in 1999. Radiocarbon dating places this individual between 7035–6680 cal BCE (93%) or 6670–6650 cal BCE (2%). Sk.21855 (burial feature 8214, Level South K, Early period, Building 17): the primary burial of a female child. She was 4 years ± 1yr at death based on dental development. It was placed in a flexed position in a burial cut along the west wall of B.17. The burial was excavated in 2016.” ref
“Sk.1885 (burial feature 84, Level South M, Middle period, Building 50): the primary flexed burial of a male child. He was 7 years ± 2yrs at death, excavated in 1995. This individual was interred directly above Sk.2033 (see below) in the southwest corner of B.50. Radiocarbon dating places this individual between 6905–6885 cal BCE (1%) or 6825–6635 cal BCE (92%) or 6625–6600 cal BCE (2%). Sk.2033 (burial feature 84, Level South M, Middle period, Building 50): the primary flexed burial of a male child 3 years ± 1yr at death, excavated in 1995. This individual was interred directly below Sk.1885 (see previous) in the southwest corner of B.50. Radiocarbon dating places this individual between 6690-6590 cal BCE (95%).” ref
“Sk.2017 (burial feature 96, Level South M, Middle period, Building 50): the primary burial of a female neonate (0-2 months at death based on measurements of the basi-occipital bone), excavated in 1997. The burial was located near the oven along the southern wall of B.50. The bones of this individual were scorched as a result of the burial’s proximity to the oven. Radiocarbon dating places this individual between 6815–6790 cal BCE (2%) or 6775–6595 cal BCE (93%) (Table Z2). Sk.2728 (burial feature 258, Level South M, Middle period, Building 50): an undisturbed primary burial of a female infant aged 9 months (±3 months) at death based on dental development. It was excavated in 1997 from Building 50, located in the South Area of the site. The body was placed in a small pit near the eastern wall of the main room. Radiocarbon dating of the petrous bone places this individual between 6695-6505 cal BCE (95%).” ref
“Sk.2779.1 (burial feature 265, Level South M, Middle period, Building 50): the primary burial of a male neonate (0-2 months at death based on measurements of the basi-occipital bone), excavated in 1997. The burial was heavily disturbed by Mellaart’s earlier excavations in this building during the 1960s. Sk.2842 (burial feature 274, Level South M, Middle period, Building 50): a disturbed primary burial of a female infant aged 18 months (±6 months) at death based on dental development. It was excavated in 1998 from Building 50, located in the South Area of the site. The body was placed in a small pit near the center of the main room and was partially disturbed by a later burial. Radiocarbon dating of the petrous bone places this individual between 6690-6505 cal BCE (95%).” ref
“Sk.21981 (burial feature 8153, Level South N, Middle period, Building 89): a disturbed primary burial of a female infant/child aged 3 years (±1 year) at death based on dental development. It was excavated in 2015 from Building 89, located in the South Area of the site. The body was placed in a small pit within the north platform of the main room and was subsequently truncated by the digging of a post retrieval pit. Sk.5747 (burial feature 1064, Level South M, Middle period, Building 91): a primary burial of a female infant aged 18 months (±6 months) at death based on dental development. It was excavated in 2002 from Building 91, located in the South Area of the site. The body was placed in a small pit located in the northeast corner of B.91. Radiocarbon dating of the petrous bone places this individual between 6640-6490 cal BCE (95%) (Table Z2).” ref
“Sk.30006 (burial feature 7615, Level North G, Middle period, Building 114): a primary burial of a female infant aged 9 months (±3 months) at death based on dental development. It was excavated in 2015. The body was interred with a middle adult female in an oval pit along the south wall of the main room. Radiocarbon dating of the petrous bone places this individual between 6645–6495 cal BCE (94%) or 6490–6480 cal BCE (1%). Sk.8587 (burial feature 1013), Level North G, Middle period, Building 114): a primary burial of a female neonate (0-2 months at death – based on long bone length) excavated in 2002 and located under the southeast platform. The burial was partially disturbed by subsequent burials in this location, and likely also by rodent burrowing.” ref
“Sk.11739 (burial feature 1912, Level TP Q-R, Final period): a heavily disturbed set of human remains belonging to a middle adult (35-50 years of age-at-death) based on dental occlusal wear. The individual was assessed as a possible male based on cranial morphology, although aDNA suggested the individual was genetically female. These remains, potentially representing a secondary burial, were excavated in 2005 from Space 411, located in the TP Area of the site. Radiocarbon dating of the petrous bone places this individual between 6235-6075 cal BCE (95%).” ref
“Sk.20217 (burial feature 3931, Level TP Q-R, Final period?): a female child aged 6 years (±2 years) at death based on dental development. This individual, excavated in 2012, is one of three individuals recovered from burial feature 3931 in the TPC Area. The burial was badly damaged as it was found directly beneath the surface. Hence, it could not be associated with any Neolithic buildings or spaces. Its stratigraphic position indicates that it post-dates B.122 from the Late period, which implies it most likely comes from the Final period. However, this is not corroborated by radiocarbon dating of the petrous bone that places this individual significantly earlier, between 6415-6240 cal BCE (95%).” ref
“ZHAJ (Area H, Grave 27): this is a primary single inhumation of a middle adult female (as determined by aDNA) buried in a sub-oval cut. The individual was found lying tightly flexed on her left side, positioned east-west with the head toward the west and facing north. ZHAG and ZHAF (Area H, Grave 18): grave 18 contained a double inhumation of a middle adult female (ZHAF) and a perinatal infant (ZHAG) found in an oval cut larger than average. The adult (ZHAF) was found lying tightly flexed on her left side and positioned with a northwest-southeast orientation with the head toward the northwest. The perinate was articulated and found with the head on top of the adult pelvis.” ref
“The female sex of the adult could be confirmed by ancient DNA. The sex of the perinate was determined as a female by aDNA, and it can be ruled out that ZHAF and ZHAG were first or second-degree related. Skeleton ZHAF has been radiocarbon dated to 8285–8175 cal BCE (83%) or 8115–8090 cal BCE (4%) or 8040–8010 cal BCE (8%). ZHB (Area H, Grave 9): a single inhumation of an adolescent-young adult female. The sex of the individual has been confirmed by ancient DNA analysis. The individual was found lying on her right side/partially prone, in a semi-flexed position. The body was orientated east-west with head to the east and facing northeast, and has been radiocarbon dated to between 8280–8165 cal BCE (57%) or 8120–7960 cal BCE (38%).” ref
“ZHF (Area H, Grave 14): single inhumation of an adult male buried in a sub-oval cut. The age-at-death of the individual was difficult to estimate accurately because both the skull and pelvis were highly fragmented. The sex has been confirmed by ancient DNA. The body was found lying on the left side and orientated northwest-southeast with the head orientated toward the northwest and facing northeast. The upper limbs were flexed at the elbow with the palms of the hands together and placed immediately in front of the face. The long bones were highly fragmented and animal burrowing had destroyed much of the skull, most of the axial elements and the feet. The skeleton has been radiocarbon dated to 8225–7940 cal BCE (95%).” ref
“ZHJ (Area H, Grave 15): this is a primary single old adult inhumation found in a sub-oval cut. The individual was found in a flexed position lying on its right side and positioned north-south with the head orientated toward the south. The bones were relatively well preserved compared with other graves, although burrowing animals had destroyed parts of the skull and axial skeleton, including the left foot. Morphological sex determination was difficult because the remains were gracile, probably as a result of the aging process. Ancient DNA analyses demonstrated that this individual was female. She has been radiocarbon dated to 8295–8240 cal BCE (95%).” ref
“ZHBJ (Area H Grave 30): single inhumation of a middle/old adult male in a suboval cut. Sex has been confirmed by ancient DNA. The individual was found lying tightly flexed on his right side, although it should be noted that there was considerable damage from bioturbation that disturbed much of the skeleton and destroyed most of the thorax and skull. The body was positioned east-west with the head toward the west, but the facing direction could not be ascertained due to the aforementioned disturbance. ZKO (Area K, Grave 12): this is a single inhumation of an old adult male in an oval cut. The individual was found lying tightly flexed on his left side and orientated east-west with the head toward the east. The bones were generally well preserved, but rodent burrowing activity caused significant disturbance of the ribs, scapulae and vertebrae. Sex was confirmed through aDNA analysis as male.” ref
“ZMOJ (Area M, Grave 49): a primary but heavily disturbed burial of a young adult male (determined by aDNA) in a sub-circular grave. The individual was orientated east-west with head to the west and facing north. The skull was found at one end of the grave and many of the other bones had been moved by animal action, so their anatomical position was not maintained. Ancient DNA indicates that this individual was male.” ref
Aşıklı Höyük
“Aşıklı Höyük is a settlement mound located nearly 1 kilometre (0.62 mi) south of Kızılkaya village on the bank of the Melendiz brook, and 25 km (16 mi) southeast of Aksaray, Turkey. Aşıklı Höyük is located in an area covered by the volcanic tuff of central Cappadocia, in Aksaray Province. The archaeological site of Aşıklı Höyük was first settled in the Pre-Pottery Neolithic period, around 8,200 BCE. It is situated 1,119.5 metres (3,673 ft) above sea level, a little higher than the region’s average of c. 1,000 metres (3,300 ft). The site itself is about 4 hectares (9.9 acres), considerably smaller than the closely situated site of Çatalhöyük (13 ha (32 acres)). The surrounding landscape is formed by erosion of river valleys into tuff deposits. The Melendiz Valley, where the Aşıklı Höyük is located, constitutes a favourable, fertile, and diverse habitat. The proximity to an obsidian source did become the base of a trade with the material supplying areas as far away as today’s Cyprus and Iraq.” ref
“Aşıklı Höyük was first investigated by Professor Ian A. Todd when he visited the site in the summer of 1964. Todd emphasised the importance of the obsidian in the area, based on over 6,000 obsidian pieces collected from the surface layer alone. The site was classified as a medium sized mound and partly destroyed by the river situated next to it. On the basis of the lithics and animal bones located in the surface layers the site became known as a contemporary to the Palestine PPNB, which later was reinforced by 14C dates (based on five unstratified radiocarbon dates going from 7008 ± 130 to 6661 ± 108). The first comprehensive excavations took place relatively late: first when the government launched a plan that would result in the rise of the waters of the Mamasın Lake located close to Aşıklı Höyük, Professor Ufuk Esin (University of Istanbul) started the salvage excavations in 1989. Nine excavations have been undertaken up to 2003, uncovering approximately 4,200 square metres (45,000 sq ft) on the horizontal plain, making it one of the largest scale excavations in the region.” ref
“The newest dates for Aşıklı Höyük show that the occupational period was from 8200 to 7400 BCE or around 10,200 to 9,400 years ago, extracted from 3 layers with a total of 13 phases; which places it in phase ECA II (correlating with the E/MPPNB in the Levant). It is known as one of the earliest Aceramic Neolithic sites on the Anatolian plateau, and the prior mentioned extraction of the obsidian source was likely to be frequented as far back as the Paleolithic nomadic hunter-gatherers. Due to its date and structural organization Aşıklı Höyük is known to be “a prime example of a first foray into sedentism.” ref
“After more than 400 rooms had been excavated, the total number of individual found to have been buried within the settlement did not surpass 70. All these burials were under building floors. The dead were placed in pits cut through the floor during the occupation of the building. The buried are people of both sexes and all ages. There is a variety of skeletal body postures, from burials in a hocker (fetal) position to extended skeletons facing upwards. Others are lying on one side, occasionally with the legs bent at the knees. The orientation of the burials varies within the buildings, as does the number of individuals buried inside them.” ref
“The male population had individuals up to the age of 55–57 years of age, while the majority of females died between the ages of 20 and 25. The skeletal remains of these women show spinal deformities indicating that they had to carry heavy loads. This does not itself prove that there was a division of labour between the sexes. The fact that the men seem to have outlived the women might be interpreted as sign that the women were subject to more strenuous physical labour than their male counterparts. From Natufian Abu Hureyra there are similar osteological signs, such as pathologies in metatarsals, phalanges, arm, and shoulder joints, being specific to females resulting from habitual kneeling in the use of saddle querns (grinding stones). The Neolithic evidence show indications of increased physical workload in the osteological material on both genders, where the male skeletons show signs of joint disease and trauma arguably caused by cutting timber and tilling.” ref
“Children represent 37.8% of the deceased, with 43.7% mortality within a year of birth. The skeletal remains are complete and with articulations intact, indicating that the burials have been primary. The graves contain either single or double burials. On one occasion two graves were found under the floor of room AB, belonging to an adjacent court (HG) with a large domed mudbrick oven paved with blocks of basalt. In one of the graves were the skeletons of a young woman and an elderly man; in the other a young woman buried together with her baby. The young woman had apparently undergone trepanation and survived only a few days after the operation.” ref
“All skeletons were buried in the hocker position, a fetal-like positioning were the arms are embracing the lower limbs. From a different grave a woman shows signs of being scalped immediately after her death, according to the cut marks on her skull. As many as 55% of the skeletons show signs of being burned. The burial under the floor AB is accommodated by walls with the interior side were painted in a purplish red colour. The oven in HG indicates that this was indeed “special individuals of an elite class”, claiming it can be compared to the “Terrazzo” Building at Çayönü and the “Temple” Building at Nevalı Çori and therefore have been a shrine used for religious ceremonies. Many of the burials contain burial goods consisting of necklaces and bracelets made of beads of various sorts.” ref
“70 burials in over 400 rooms suggest that some form of selection took place of who was buried at the site, implying that AB indeed could be the residence or resting place of people influential in terms of both economy and political power. Rooms containing hearths are more likely to contain burials; as many as 77%. It has been argued that the number of burials could be an underrepresentation inhered at the site, since a large part of the settlement remain unexcavated beneath the baulks. Later excavations which have been published suggest on the other hand that burials were not a general feature at Aşıklı Höyük and therefore the suggestions of burials being a privilege of the elite class do seem plausible. There has not been found a cemetery or any other sign of where the rest of the population might have been disposed of post mortem. This issue is not only limited to Aşıklı Höyük: there is also a lack of cemeteries on the PPNB “mega-sites” in the Levant, such ‘Ain Ghazal in the Jordan Valley.” ref
“It seems that in Aşıklı Höyük, as in the rest of the Anatolian and Levantine area, the burial and any other post mortem treatment was arguably an “upper class” phenomenon. This interpretation has been opposed, referring to the diversity of individuals in both sex and age in the graves. The burials including such a wide range of individuals do not directly coherent with the image of an “upper class” phenomenon. Burials could have been removed or replaced over time, giving a wrong image of the burials as belonging to the elite. An alternative perspective has been suggested: “the Neolithic dead are not under-represented: rather, it is the architecture in settlements that are over-represented”, meaning that in many cases archaeologists have drastically overestimated the extent to which all areas of Neolithic sites were occupied simultaneously. As for Aşıklı Höyük and other sites in the area: low numbers of burials in comparison with occupation span does not directly indicate a cult of the elite.” ref
“At Aşıklı Höyük the hearths are rectangular and usually placed in one of the corners of the rooms, ranging in size from 2.97 to 0.48 square metres (32.0 to 5.2 sq ft). Large stones with a suitable flat shape were used to create an upright edge that stood approximately 20 cm (7.9 in) above the level of the floor. On the short side of the hearth the upright edge is missing to make a fire mouth. It is also here the ash is the most concentrated. Pebbles along the edges and base of the hearth seem to have been covered by a thin layer of plaster. Only in a few cases there is a trace of something recognized as a flue. An estimated 30–40% of all the rooms at Aşıklı Höyük have hearths. This estimate is based on partially damaged and eroded structures possibly giving a number lower than accurate. Based on a limited group of fully exposed buildings 54% of single room units contained a hearth, while only 29% of the multiple room units. The average percentage on base of these building units is 47%, probably a more realistic estimate for the site in total.” ref
“Hearths do not occur in a courtyard context, and are more represented in single room dwellings than multiple room units. Still, multiple room units do have a substantial number of hearths. It has been suggested that the “multiroom dwellings may have functioned as ‘incomplete’ houses for new families still heavily dependent on the larger extended group”. The buildings containing the hearths do not show particular characteristics that distinguish them from structures without hearths; neither do they differ in size or special orientation. Even the hearth itself does not follow an apparent norm in terms of size or location. The position varies considerably, but it always has one side to the wall. The positioning of the hearth does not seem to be determined by general macro-ecological factors, such as prevailing wind directions, nor determined by cultural norms regarding spatial features within buildings.” ref
“The hearth does not seem to be subject to a random placement inside the buildings: it is consistently located at the same spot throughout a very long building sequence. This indicates that their positions were not chosen arbitrarily. When a location for the hearth was chosen it was important that the placement did not change in later rebuilding sequences of the structure (see picture: Deep sounding). There is no evidence for ladderpost scars due to the assumed use of freestanding ladders, making the location of the entrance uncertain. Aşıklı Höyük does not seem to have any evidence for ovens. Aşıklı Höyük had a tradition to reconstruct or rebuild earlier structures. It followed a pattern where the structures were built “exactly on the same spot and with the same alignment as earlier buildings, using older walls as a foundation.” ref
“The structural continuity at Aşıklı Höyük is outstanding, but there is no information to how long the use-life of a building was. If one estimates the same lifespan for a structure at Aşıklı Höyük as it was in Çatalhöyük, one could look at an age of 30 to 60 years before reconstruction occurred. If this assumption is correct, the deep sounding 4H/G from phases 2I up to 2B (eight layers in total) show that the time span of a structure could be from 240 to 480 years. Looking outside of Central Anatolian Neolithic, this type of building continuity is unparalleled both in ethnography and archaeology. This remarkable structural continuity may suggest a social system in which buildings were not privately owned, since one would expect them to be modified on a regular basis. It can be assumed that the rooms were distributed amongst the community members according to the change in both needs and statuses.” ref
“The building practices maintained their characteristics throughout the centuries. It has been claimed that the building continuity is a self-evident feature, since it is deriving from a particular set of foundation practices that can be explained in a functionalistic way. It has otherwise been argued that the extreme degree of continuity is inadequately explained by functionalism alone, since the structures located adjacent to open spaces could have easily been expanded or shrunk according to the specific needs, but instead remained identical. These functionalist parameters can also not explain the continued rebuilding of the hearths, which are always built on the same spot. Individual hearth sequences are often separated with 40 cm (16 in) of soil, and therefore there is no apparent reason (unlike the buildings) why the hearths should consistently be constructed in the same corner as in the successive buildings. In many cases neighbouring buildings do place their respective hearths in different corners. Micro-cosmological special codes or wind direction does not seem to be decisive for the positioning of the hearth. The structural and material remains indicate that the buildings were continuous entities with some form of fixed special identity where the special organization could not be changed by the temporary occupant.” ref
“Structural continuity was of great importance to the inhabitants of Aşıklı Höyük. The reason for this has partly been explained because they (the people) had a rigid adherence to traditions in terms of structural reproduction. The “traditional view” has been disputed because “[In short,] labelling a society as conservative does not answer the question why the people under consideration were conservative”. An alternative approach refers to the historical dimension of the building to be of such great importance that “people came to be bound between walls, metaphorically domesticated”. The interpretation is that the walls are giving historical associations to the people living within them, giving a collective conscience lasting through time. The difference between this interpretation and the “conservative approach” is the potential explanation to why structural reproduction could have been important for the inhabitants of Aşıklı Höyük. The identities of the inhabitants were projected to the structural outcome of the buildings. The generality of this position is not meaningful on its own. It is not certain that the inhabitants of Aşıklı Höyük were aware of the total amount of building there was in the sequence in total. Perhaps the history of a building did not concern them in the same way as archaeologists like to think.” ref
“The lack of change over time suggests that the inhabitants of Aşıklı Höyük had a view of the past as a precedent for the present: a vital part of society that was ‘reborn’ in each reproduction, manifested in its building continuity. The structural reconstruction is a regional feature for Central Anatolia. With the exception of Jericho, most of the evidence from PPNB sites in the Levant indicates that structures were not reconstructed in the same loci, and some location structures differ in dates by several hundred years. The buildings at Aşıklı Höyük are clustered into what has been interpreted as neighbourhoods. As this is a vague perception of the structural outlay of the community it describes them as clustered single and multiroom houses forming compounds, apparently sharing courtyard space for production activities and practising joint cooking and food consumption. Little can be said on the food storage, since there were no remains after storage bins, although storage rooms may be identified due to comparing structures on other sites (e.g. Çatalhöyük).” ref
“The average room size is 12 m2 (130 sq ft) (at this time humans were 1.5 to 1.7 metres (4 ft 11 in to 5 ft 7 in) tall). From two or three up to five or six clustered dwellings formed a ‘neighborhood’ or compound. The interpretation of the borders of these ‘neighbourhoods’ is problematic, since much of the site still lies under the baulks, is in situ or eroded. The distribution of single- and multi-room buildings does not seem to follow a pattern other than that the residential clusters seem to be divided by narrow alleys 0.5–1.0 m (1 ft 8 in – 3 ft 3 in) wide, or open courtyard areas up to a diameter of 4 m (13 ft). The interior of multiroom buildings had openings in the partitioned walls, providing access to the individual rooms. Between the one building and the next there seems to be no communication, since there was no indication of doors in the exterior mudbrick walls. Since the buildings themselves do not have an entrance that can be traced archaeologically on the base of the walls, access had to be provided either through window-like openings high on the walls or from the flat roofs. Roof access is also known from Çatalhöyük, making this entrance more plausible.” ref
“Aşıklı Höyük does also have buildings that are bigger in size but without hearths. These are interpreted as public buildings or ‘building complexes’. These are seen as some of the most enigmatic buildings found at the site, and diverge both in size and spatial organization. One of them (complex HV) being at up to 20 times larger than the largest loam buildings (i.e. 25 m × 20 m (82 ft × 66 ft) = 500 m2 (5,400 sq ft)). They have a multitude of rooms and encompass elaborate and large internal courts; something that is not found in any other buildings. The walls are more robust and massive than other buildings, in some cases being referred to as “monumental walls”, accompanied by parallel outer walls with relatively narrow space in between.” ref
“The interpretation of these buildings is difficult. The fact that they clearly differentiate from the domestic loam buildings indicates that they had special value in the society. They also do not incorporated into the clustered ‘neighbourhoods’, indicating that they served several neighbourhoods or the local community at large. With 500 m2 (5,400 sq ft) the range of activities that could have taken place in this space could easily incorporate several hundred people. Yet, given that the estimated population of Aşıklı Höyük may have run into the thousands, only a selected group in the total population could have used the building at a given occasion. There is a variety of hypotheses regarding the nature of these monumental structures. There are other examples of these restricted monumental spaces on other sites in the Levantine PPNB (such as Nevali Çori, Behida, ‘Ain Ghazal), suggesting that they were used by an elite or for practising different social initiation rites.” ref
“There are no finds of any artefacts carrying religious connotations, symbolic or imagery, at Aşıklı Höyük, in the buildings, courtyards, dumps or open-workshop areas. The only finds include flint tools, which are counted as imports. Other than this there is found one single animal figurine made of clay that can hardly tell us anything of the religious belief of the inhabitants. The limited amount of burials compared to the estimated population makes it very likely that there may have been a cemetery where the deceased were buried, but it has not been found. There is also an absence of storage bins, making the distinction on autonomous households difficult.” ref
Samara, Russia 7,500 years ago carried R1b1a1a
“An EHG buried near Samara, Russia 7,500 years ago carried R1b1a1a. An Eneolithic male buried at Khvalynsk, Russia 7,200-6,000 years ago carried R1b1a.” ref
R1a was not found in Yamnaya remains
“The most common Y-DNA haplogroup found among the Yamnaya specimens was Haplogroup R1b, specifically the Z2103 subclade of R1b-L23. Additionally, a minority are found to belong to haplogroup I2. Autosomal tests also indicate that the Yamnaya are the vector for “Ancient North Eurasian” admixture into Europe. “Ancient North Eurasian” is the name given in literature to a genetic component that represents descent from the people of the Mal’ta–Buret’ culture or a population closely related to them. That genetic component is visible in tests of the Yamnaya people as well as modern-day Europeans.” ref, ref, ref
“Mascarenhas et al. (2015) proposed that the roots of Z93 lie in West Asia, and proposed that “Z93 and L342.2 expanded in a southeasterly direction from Transcaucasia into South Asia“, noting that such an expansion is compatible with “the archeological records of eastward expansion of West Asian populations in the 4th millennium BCE culminating in the so-called Kura-Araxes

Kunda culture
“The earliest traces of human settlement in Estonia are connected with the Kunda culture. The early Mesolithic Pulli settlement is located by the Pärnu River. It has been dated to the beginning of the 9th millennium BCE. The Kunda culture received its name from the Lammasmäe settlement site in northern Estonia, which dates from earlier than 8500 BCE. Bone and stone artifacts similar to those found at Kunda have been discovered elsewhere in Estonia, as well as in Latvia, northern Lithuania, and southern Finland. Among minerals, flint and quartz were used the most for making cutting tools. The beginning of the Neolithic Period is marked by the ceramics of the Narva culture, which appeared in Estonia at the beginning of the 5th millennium BCE. The oldest finds date from around 4900 BCE.” ref
“The Kunda culture, which originated from the Swiderian culture, comprised Mesolithic hunter-gatherer communities of the Baltic forest zone extending eastwards through Latvia into northern Russia, dating to the period 8500–5000 BCE according to calibrated radiocarbon dating. It is named after the Estonian town of Kunda, about 110 kilometres (70 mi) east of Tallinn along the Gulf of Finland, near where the first extensively studied settlement was discovered on Lammasmäe Hill and in the surrounding peat bog. The oldest known settlement of the Kunda culture in Estonia is Pulli. The Kunda culture was succeeded by the Narva culture, who used pottery and showed some traces of food production.” ref
“Most Kunda settlements are located near the edge of the forests beside rivers, lakes, or marshes. Elk were extensively hunted, perhaps helped by trained domestic hunting-dogs. On the coast seal hunting is represented. Pike and other fish were taken from the rivers. There is a rich bone and antler industry, especially in relation to fishing gear. Tools were decorated with simple geometric designs, lacking the complexity of the contemporary Maglemosian Culture communities to the southwest. Jones et al. (2017) determined, based on one sample (6467-6250 BCE) from the Kunda culture and another one from the succeeding Narva culture, closer genetic affinity with Western Hunter-Gatherers (WHGs) than Eastern Hunter-Gatherers (EHGs).” ref
“The Kunda culture appears to have undergone a transition from the Palaeolithic Swiderian culture located previously over much of the same range. One such transition settlement, Pasieniai 1C in Lithuania, features stone tools of both Late Swiderian and early Kunda. One shape manufactured in both cultures is the retouched tanged point. The final Swiderian is dated 7800–7600 BCE by calibrated radiocarbon dating, which is in the Preboreal period, at the end of which time with no gap the early Kunda begins. Evidently, the descendants of the Swiderians were the first to settle Estonia when it became habitable. Other post-Swiderian groups extended as far east as the Ural Mountains. Mittnik et al. (2018) analyzed the remains of a male and female ascribed to the Kunda culture. They found the male to be carrying paternal haplogroup I and maternal haplogroup U5b2c1, while the female carried U4a2.” ref
“They were found to have “a very close affinity” with WHGs, although with “a significant contribution” from Ancient North Eurasians (ANE). Their ANE ancestry was lower than that of Scandinavian Hunter-Gatherers, indicating that ANE ancestry entered Scandinavia without traversing the Baltic. Matthieson et al. (2018) analyzed a large number of individuals buried at the Zvejnieki burial ground, most of whom were affiliated with the Kunda culture and the succeeding Narva culture. The mtDNA belonged to haplotypes U5, U4 and U2, the vast majority of the Y-DNA samples belonged to R1b1a1a and I2a1. The results affirmed that the Kunda and Narva cultures were about 70% WHG and 30% EHG. The nearby contemporary Pit–Comb Ware culture was on the contrary found to be about 65% EHG. Around 3700/3600 BC there was a complete turnover to now y-hg R1a1a1, from 900/800 onward mainly in Estonia with increasing impact of y-hg N-M231 (N1a1).” ref
“The first pottery was made of thick clay mixed with pebbles, shells, or plants. The Narva-type ceramics are found throughout almost the entire Estonian coastal region and on the islands. The stone and bone tools of the era have a notable similarity with the artifacts of the Kunda culture. Around the beginning of the 4th millennium BCE, Comb Ceramic culture arrived in Estonia. Until the early 1980s, the arrival of Finnic peoples, the ancestors of the Estonians, Finns, and Livonians, on the shores of the Baltic Sea was associated with the Comb Ceramic Culture. However, such a linking of archaeologically defined cultural entities with linguistic ones cannot be proven, and it has been suggested that the increase of settlement finds in the period is more likely to have been associated with an economic boom related to the warming of the climate. Some researchers have even argued that a Uralic form of language may have been spoken in Estonia and Finland since the end of the last glaciation.” ref
“The burial customs of the comb pottery people included additions of figures of animals, birds, snakes, and men carved from bone and amber. Antiquities from comb pottery culture are found from Northern Finland to Eastern Prussia. The beginning of the Late Neolithic Period, about 2200 BCE, is characterized by the appearance of the Corded Ware culture, pottery with corded decoration, and well-polished stone axes (s.c. boat-shape axes). Evidence of agriculture is provided by charred grains of wheat on the wall of a corded-ware vessel found in the Iru settlement. Osteological analysis shows an attempt was made to domesticate the wild boar. Specific burial customs were characterized by the dead being laid on their sides with their knees pressed against their breast, one hand under the head. Objects placed into the graves were made of the bones of domesticated animals.” ref
“The beginning of the Bronze Age in Estonia is dated to approximately 1800 BCE. The development of the borders between the Finnic peoples and the Balts was under way. The first fortified settlements, Asva and Ridala on the islands of Saaremaa and Iru in Northern Estonia, began to be built. The development of shipbuilding facilitated the spread of bronze. Changes took place in burial customs, a new type of burial ground spread from Germanic to Estonian areas, and stone cist graves and cremation burials became increasingly common, alongside a small number of boat-shaped stone graves. About the 7th century BCE or around 2,700 years ago, a large meteorite hit Saaremaa island and created the Kaali craters. About 325 BCE, the Greek explorer Pytheas possibly visited Estonia. The Thule island he described has been identified as Saaremaa by Lennart Meri, though this identification is not widely considered probable, as Saaremaa lies far south of the Arctic Circle.” ref
“The Pre-Roman Iron Age began in Estonia about 500 BC and lasted until the middle of the 1st century CE. The oldest iron items were imported, although since the 1st century iron was smelted from local marsh and lake ore. Settlement sites were located mostly in places that offered natural protection. Fortresses were built, although used temporarily. The appearance of square Celtic fields surrounded by enclosures in Estonia date from the Pre-Roman Iron Age. The majority of stones with man-made indents, which presumably were connected with magic designed to increase crop fertility, date from this period. A new type of grave, quadrangular burial mounds, began to develop. Burial traditions show the clear beginning of social stratification.” ref
Hunter-Gatherer Pottery?
“Pottery is one of the oldest human inventions, originating before the Neolithic period, with ceramic objects such as the Gravettian culture Venus of Dolní Věstonice figurine discovered in the Czech Republic dating back to 29,000–25,000 BCE. However, the earliest known pottery vessels were discovered in Jiangxi, China, which date back to 18,000 BC. Other early Neolithic and pre-Neolithic pottery artifacts have been found, in Jōmon Japan (10,500 BCE), the Russian Far East (14,000 BCE), Sub-Saharan Africa (9,400 BCE), South America (9,000s–7,000s BCE), and the Middle East (7,000s–6,000s BCE).” ref
“The earliest forms of pottery were made from clays that were fired at low temperatures, initially in pit-fires or in open bonfires. They were hand-formed and undecorated. Earthenware can be fired as low as 600 °C, and is normally fired below 1200 °C. Because unglazed earthenware is porous, it has limited utility for the storage of liquids or as tableware. However, earthenware has had a continuous history from the Neolithic period to today. It can be made from a wide variety of clays, some of which fire to a buff, brown or black colour, with iron in the constituent minerals resulting in a reddish-brown. Reddish coloured varieties are called terracotta, especially when unglazed or used for sculpture. The development of ceramic glaze made impermeable pottery possible, improving the popularity and practicality of pottery vessels. Decoration has evolved and developed through history.” ref
“Stoneware is pottery that has been fired in a kiln at a relatively high temperature, from about 1,100 °C to 1,200 °C, and is stronger and non-porous to liquids. The Chinese, who developed stoneware very early on, classify this together with porcelain as high-fired wares. In contrast, stoneware could only be produced in Europe from the late Middle Ages, as European kilns were less efficient, and the right type of clay less common. It remained a speciality of Germany until the Renaissance. Stoneware is very tough and practical, and much of it has always been utilitarian, for the kitchen or storage rather than the table. But “fine” stoneware has been important in China, Japan and the West, and continues to be made. Many utilitarian types have also come to be appreciated as art.” ref
“The study of pottery can help to provide an insight into past cultures. Fabric analysis (see section below), used to analyze the fabric of pottery, is an important part of archaeology for understanding the archaeological culture of the excavated site by studying the fabric of artifacts, such as their usage, source material composition, decorative pattern, color of patterns, etc. This helps to understand the characteristics, sophistication, habits, technology, tools, trade, etc., of the people who made and used the pottery. Carbon dating reveals the age. Sites with similar pottery characteristics have the same culture; those sites that have distinct cultural characteristics but with some overlap are indicative of cultural exchange, such as trade or living in the vicinity or continuity of habitation, etc. Examples are black and red ware, redware, Sothi-Siswal culture, and Painted Grey Ware culture. The six fabrics of Kalibangan is a good example of use of fabric analysis in identifying a differentiated culture which was earlier thought to be typical Indus Valley civilisation (IVC) culture.” ref
“Pottery is durable, and fragments, at least, often survive long after artifacts made from less-durable materials have decayed past recognition. Combined with other evidence, the study of pottery artifacts is helpful in the development of theories on the organization, economic condition, and cultural development of the societies that produced or acquired pottery. The study of pottery may also allow inferences to be drawn about a culture’s daily life, religion, social relationships, attitudes towards neighbors, attitudes to their own world, and even the way the culture understood the universe. It is valuable to look into pottery as an archaeological record of potential interaction between peoples. When pottery is placed within the context of linguistic and migratory patterns, it becomes an even more prevalent category of social artifact.” ref
“ Sherds have been found in China and Japan from a period between 12,000 and perhaps as long as 18,000 years ago. As of 2012, the earliest pottery vessels found anywhere in the world, dating to 20,000 to 19,000 years before the present, was found at Xianren Cave in the Jiangxi province of China. Other early pottery vessels include those excavated from the Yuchanyan Cave in southern China, dated from 16,000 BCE, and those found in the Amur River basin in the Russian Far East, dated from 14,000 BCE. The Odai Yamamoto I site, belonging to the Jōmon period, currently has the oldest pottery in Japan. Excavations in 1998 uncovered earthenware fragments which have been dated as early as 14,500 BCE. The term “Jōmon” means “cord-marked” in Japanese. This refers to the markings made on the vessels and figures using sticks with cords during their production. Recent research has elucidated how Jōmon pottery was used by its creators.” ref
“It appears that pottery was independently developed in Sub-Saharan Africa during the 10th millennium BCE, with findings dating to at least 9,400 BCE from central Mali, and in South America during the 9,000s–7,000s BCE. The Malian finds date to the same period as similar finds from East Asia – the triangle between Siberia, China and Japan – and are associated in both regions to the same climatic changes (at the end of the ice age new grassland develops, enabling hunter-gatherers to expand their habitat), met independently by both cultures with similar developments: the creation of pottery for the storage of wild cereals (pearl millet), and that of small arrowheads for hunting small game typical of grassland. Alternatively, the creation of pottery in the case of the Incipient Jōmon civilisation could be due to the intensive exploitation of freshwater and marine organisms by late glacial foragers, who started developing ceramic containers for their catch.” ref
“Around 8000 BCE during the Pre-pottery Neolithic period, and before the invention of pottery, several early settlements became experts in crafting beautiful and highly sophisticated containers from stone, using materials such as alabaster or granite, and employing sand to shape and polish. Artisans used the veins in the material to maximum visual effect. Such objects have been found in abundance on the upper Euphrates river, in what is today eastern Syria, especially at the site of Bouqras. The earliest history of pottery production in the Fertile Crescent starts the Pottery Neolithic and can be divided into four periods, namely: the Hassuna period (7000–6500 BCE), the Halaf period (6500–5500 BCE), the Ubaid period (5500–4000 BCE), and the Uruk period (4000–3100 BCE). By about 5000 BCE pottery-making was becoming widespread across the region, and spreading out from it to neighbouring areas. Pottery making began in the 7th millennium BC. The earliest forms, which were found at the Hassuna site, were hand formed from slabs, undecorated, unglazed low-fired pots made from reddish-brown clays.” ref
“Within the next millennium, wares were decorated with elaborate painted designs and natural forms, incising and burnished. The invention of the potter’s wheel in Mesopotamia sometime between 6,000 and 4,000 BC (Ubaid period) revolutionised pottery production. Newer kiln designs could fire wares to 1,050 °C (1,920 °F) to 1,200 °C (2,190 °F) which enabled increased possibilities. Production was now carried out by small groups of potters for small cities, rather than individuals making wares for a family. The shapes and range of uses for ceramics and pottery expanded beyond simple vessels to store and carry to specialized cooking utensils, pot stands and rat traps. As the region developed new organizations and political forms, pottery became more elaborate and varied. In the Chalcolithic period in Mesopotamia, Halafian pottery achieved a level of technical competence and sophistication, not seen until the later developments of Greek pottery with Corinthian and Attic ware.” ref
“Europe’s oldest pottery, dating from circa 6700 BCE, was found on the banks of the Samara River in the middle Volga region of Russia. These sites are known as the Yelshanka culture. The early inhabitants of Europe developed pottery in the Linear Pottery culture slightly later than the Near East, circa 5500–4500 BCE. In the ancient Western Mediterranean elaborately painted earthenware reached very high levels of artistic achievement in the Greek world; there are large numbers of survivals from tombs. Minoan pottery was characterized by complex painted decoration with natural themes. The classical Greek culture began to emerge around 1000 BC featuring a variety of well crafted pottery which now included the human form as a decorating motif.” ref
“The pottery wheel was now in regular use. Although glazing was known to these potters, it was not widely used. Instead, a more porous clay slip was used for decoration. A wide range of shapes for different uses developed early and remained essentially unchanged during Greek history. Fine Etruscan pottery was heavily influenced by Greek pottery and often imported Greek potters and painters. Ancient Roman pottery made much less use of painting, but used moulded decoration, allowing industrialized production on a huge scale. Much of the so-called red Samian ware of the Early Roman Empire was produced in modern Germany and France, where entrepreneurs established large potteries.” ref
Narva culture (First European Pottery?)
“Preceded by the Kunda culture, and followed by the Pit–Comb Ware culture, Corded Ware culture, and Brushed Pottery culture. The Narva culture or eastern Baltic was a European Neolithic archaeological culture in present-day Estonia, Latvia, Lithuania, Kaliningrad Oblast (former East Prussia), and adjacent portions of Poland, Belarus, and Russia. A successor of the Mesolithic Kunda culture, the Narva culture continued up to the start of the Bronze Age. The culture spanned from c. 5300 to 1750 BCE. The technology was that of hunter-gatherers. The culture was named after the Narva River in Estonia.” ref
“The people of the Narva culture had little access to flint; therefore, they were forced to trade and conserve their flint resources. For example, there were very few flint arrowheads and flint was often reused. The Narva culture relied on local materials (bone, horn, schist). As evidence of trade, researchers found pieces of pink flint from Valdai Hills and plenty of typical Narva pottery in the territory of the Neman culture while no objects from the Neman culture were found in Narva. Heavy use of bones and horns is one of the main characteristics of the Narva culture. The bone tools, continued from the predecessor Kunda culture, provide the best evidence of continuity of the Narva culture throughout the Neolithic period. The people were buried on their backs with few grave goods. The Narva culture also used and traded amber; a few hundred items were found in Juodkrantė. One of the most famous artifacts is a ceremonial cane carved of horn as a head of female elk found in Šventoji.” ref
“The people were primarily fishers, hunters, and gatherers. They slowly began adopting husbandry in the middle Neolithic. They were not nomadic and lived in the same settlements for long periods as evidenced by abundant pottery, middens, and structures built in lakes and rivers to help fishing. The pottery shared similarities with the Comb Ceramic culture, but had specific characteristics. One of the most persistent features was mixing clay with other organic matter, most often crushed snail shells. The pottery was made of 6-to-9 cm (2.4-to-3.5 in) wide clay strips with minimal decorations around the rim. The vessels were wide and large; the height and the width were often the same. The bottoms were pointed or rounded, and only the latest examples have narrow flat bottoms. From mid-Neolithic, Narva pottery was influenced and eventually disappeared into the Corded Ware culture.” ref
“The prehistoric shell middens of Atlantic Europe consist of marine molluscs, but the eastern Baltic did not have exploitable marine species. Here the sole recorded shell midden, at Riņņukalns in Latvia, is on an inland lake and is formed of massive dumps of freshwater shells. Recent excavations indicate that they are the product of a small number of seasonal events during the later fourth millennium BCE. The thickness of the shell deposits suggests that this was a special multi-purpose residential site visited for seasonal aggregations by pottery-using hunter-gatherer communities on the northern margin of Neolithic Europe.” ref
“For a long time, archaeologists believed that the first inhabitants of the region were Finno-Ugric, who were pushed north by people of the Corded Ware culture. In 1931, Latvian archaeologist Eduards Šturms was the first to note that artifacts found near the Zebrus Lake in Latvia were different and possibly belonged to a separate archaeological culture. In early 1950s settlements on the Narva River were excavated. Lembit Jaanits and Nina Gurina grouped the findings with similar artifacts from eastern Baltic region and described the Narva culture. At first, it was believed that Narva culture ended with the appearance of the Corded Ware culture.” ref
“However, newer research extended it up to the Bronze Age. As Narva culture spanned several millennia and encompassed a large territory, archaeologists attempted to subdivide the culture into regions or periods. For example, in Lithuania two regions are distinguished: southern (under influence of the Neman culture) and western (with major settlements found in Šventoji). There is an academic debate what ethnicity the Narva culture represented: Finno-Ugrians or other Europids, preceding the arrival of the Indo-Europeans. It is also unclear how the Narva culture fits with the arrival of the Indo-Europeans (Corded Ware and Globular Amphora cultures) and the formation of the Baltic tribes.” ref
“Mathieson (2015) analyzed a large number of individuals buried at the Zvejnieki burial ground, most of whom were affiliated with the Kunda culture and the succeeding Narva culture. The mtDNA extracted belonged exclusively to haplotypes of U5, U4, and U2. With regards to Y-DNA, the vast majority of samples belonged to R1b1a1a haplotypes and I2a1 haplotypes. The results affirmed that the Kunda and Narva cultures were about 70% WHG and 30% EHG. The nearby contemporary Pit–Comb Ware culture was, on the contrary, found to be about 65% EHG. An individual from the Corded Ware culture, which would eventually succeed the Narva culture, was found to have genetic relations with the Yamnaya culture.” ref
“Jones et al. (2017) examined the remains of a male of the Narva culture buried c. 5780-5690 BCE. He was found to be a carrier of the paternal haplogroup R1b1b and the maternal haplogroup U2e1. People of the Narva culture and preceding Kunda culture were determined to have closer genetic affinity with Western Hunter-Gatherers (WHGs) than Eastern Hunter-Gatherers (EHGs). Saag et al. (2017) determined haplogroup U5a2d in a Narva male. Mittnik et al. (2018) analyzed 24 Narva individuals. Of the four samples of Y-DNA extracted, one belonged to I2a1a2a1a, one belonged to I2a1b, one belonged to I, and one belonged to R1. Of the ten samples of mtDNA extracted, eight belonged to U5 haplotypes, one belonged to U4a1, and one belonged to H11. U5 haplotypes were common among Western Hunter-Gatherers (WHGs) and Scandinavian Hunter-Gatherers (SHGs). Genetic influence from Eastern Hunter-Gatherers (EHGs) was also detected.” ref
Pitted Ware culture
“The Pitted Ware culture (c. 3500 BC–c. 2300 BCE) was a hunter-gatherer culture in southern Scandinavia, mainly along the coasts of Svealand, Götaland, Åland, north-eastern Denmark and southern Norway. Despite its Mesolithic economy, it is by convention classed as Neolithic, since it falls within the period in which farming reached Scandinavia. The Pitted Ware people were largely maritime hunters, and were engaged in lively trade with both the agricultural communities of the Scandinavian interior and other hunter-gatherers of the Baltic Sea. The people of the Pitted Ware culture were a genetically homogeneous and distinct population descended from earlier Scandinavian Hunter-Gatherers (SHGs). The culture emerged in east-central Sweden around 3,500 BC, gradually replacing the Funnelbeaker culture throughout the coastal areas of southern Scandinavia. It subsequently co-existed with the Funnelbeaker culture for several centuries.” ref
“From about 2,800 BCE, the Pitted Ware culture co-existed with the Battle Axe culture, which was the successor of the Funnelbeaker culture in southern Scandinavia. By 2,300 BCE, the Pitted Ware culture had been absorbed by the Battle Axe culture. The subsequent Nordic Bronze Age represents a fusion of elements from the Pitted Ware culture and the Battle Axe culture. Modern Scandinavians, unlike the Sami, display partial genetic origins from the Pitted Ware people. Genetic studies suggest that the Pitted Ware peoples, unlike their Neolithic neighbors, were descended from earlier Scandinavian Hunter-Gatherers (SHGs). At the time of the emergence of the Pitted Ware culture, these hunter-gatherers persisted to the north of the agricultural Funnelbeaker culture. Their ceramic traditions are related to those of the Comb Ceramic culture. Because of this, Pitted Ware Culture is often combined with Comb Ceramic Culture to form Pit Comb Ware Culture.” ref
“The Pitted Ware culture arose around 3,500 BCE. Its earliest sites are found in east-central Sweden, where it appears to have replaced the Funnelbeaker culture. Its subsequent expansion is accompanied by the disappearance of settlements of the Funnelbeaker culture throughout large parts of southern Scandinavia. It came to occupy the coasts of Denmark, southern Sweden, southern Norway and various islands of the Baltic Sea, such as Öland, Gotland, and Åland. There were lively contacts with hunter-gatherer communities of Finland and the eastern Baltic. During its initial years, the Pitted Ware culture co-existed with the Funnelbeaker culture. Although the two cultures exchanged goods with each other, its peoples appear to have had widely different identities, and they did not mix with each other to any notable extent. During the period of Pitted Ware expansion, the Funnelbeakers constructed a number of defensive palisades, which may mean that the two peoples were in conflict with each other. There is archaeological evidence of high levels of violence among the people of the Pitted Ware culture. Throughout its existence of more than 1,000 years, the Pitted Ware culture remained virtually unchanged.” ref
“From around 2,800 BCE, the Pitted Ware culture co-existed for some time with the Battle Axe culture and the Single Grave culture, which succeeded the Funnelbeaker culture in southern Scandinavia. Both were variants of the Corded Ware culture. Like the Funnelbeakers, the Corded Ware constructed a series of defensive palisades during this period, which may be a sign of violent conflict between them and the Pitted Ware. Though cultural influences of the Battle Axe culture are detectable in Pitted Ware burials, its peoples do not appear to have mixed with each other. By ca. 2,300 BCE, the Pitted Ware culture had merged with the Battle Axe culture. The subsequent Nordic Bronze Age represents a fusion of elements from the Pitted Ware culture and the Battle Axe culture.” ref
“The economy of the Pitted Ware culture was based on fishing, hunting and gathering of plants. Pitted Ware sites contain bones from elk, deer, beaver, seal, porpoise, and pig. Pig bones found in large quantities on some Pitted Ware sites emanate from wild boar rather than domestic pigs. The hunting of seal was particularly important. For this reason, the Pitted Ware people have been called “hard-core sealers” or the “Inuit of the Baltic”. Seasonal migration was a feature of life, as with many other hunter-gatherer communities. Pitted Ware communities in Eastern Sweden probably spent most of the year at their main village on the coast, making seasonal forays inland to hunt for pigs and fur-bearing animals and to engage in exchange with farming communities in the interior.” ref
“This type of seasonal interaction may explain the unique Alvastra Pile Dwelling in south-western Östergötland, which belongs to the Pitted Ware culture as far as the pottery is concerned, but to the Funnelbeaker culture in tools and weapons. The Pitted Ware peoples appear to have been specialized hunters who engaged in the trade of animal goods with peoples throughout the Baltic. The repertoire of Pitted Ware tools varied from region to region. In part this variety reflected regional sources of raw materials. However, the use of fish hooks, harpoons, nets, and sinkers was fairly widespread. Tanged arrow heads made from blades of flintstone are abundant on Scandinavia’s west coast, and were probably used in the hunting of marine mammals.” ref
“One notable feature of the Pitted Ware Culture is the sheer quantity of shards of pottery on its sites. The culture has been named after the typical ornamentation of its pottery: horizontal rows of pits pressed into the body of the pot before firing. Though some vessels are flat-bottomed, others are round-based or pointed-based, which would facilitate stable positioning in the soil or on the hearth. In shape and decoration, these ceramics reflect influences from the Comb Ceramic culture (also known as Pit-Comb Ware) of Finland and other parts of north-eastern Europe, established in the sixth and fifth millennia BCE. Small animal figurines were modelled from clay, as well as bone. These are also similar to the art of the Comb Ware culture. A large number of ceramic figurines have been found at Jettböle on the island of Jomala in Åland, including some which combine seal and human features.” ref
“The Pitted Ware people buried their dead in cemeteries. Most excavated Pitted Ware burials are located at Gotland, where around 180 graves have been found at numerous sites with several layers. One such site is at Västerbjers. Pitted Ware people were typically buried in flat inhumation graves, although cremation does occur. Unlike the Funnelbeakers, they did not have megalithic graves. Pitted Ware burials are also distinguished from Funnelbeaker burials through their use of red ochre. Grave goods include ceramics, boar tusks, pig jaws, pendants of fox, dog and seal teeth, harpoons, spears, fishhooks of bone, stone and flint axes, and other artifacts. The presence of slate artifacts and battle axes attest wide-ranging contacts between the Pitted Ware people and other cultures of Northern Europe and the Baltic. People of all ages and genders were buried in the same cemetery. There are no indications of difference in social status. Their mortuary houses and secondary burials are nevertheless evidence of complex burial customs.” ref
“The Pitted Ware people had an animistic cosmography similar to that of the people of the Comb Ceramic culture and other Mesolothic hunter-gatherers of the Baltic. Genetic studies of the Pitted Ware peoples have found them to have been strikingly genetically homogenous, suggesting that they originated from a small founder group. Already Malmström (2009) extracted mtDNA from seventeen Pitted Ware people of Gotland. Eight individuals belonged to U4 haplotypes, seven belonged to U5 haplotypes, one to K1a1, one to T2b, and one to HV0. The results debunked previous theories suggesting that the Pitted Ware were related to the Sami people. On the contrary, Pitted Ware people showed closer genetic kinship to modern Balts, Estonians, and modern Scandinavians.” ref
“In a genetic study published in BMC Evolutionary Biology in March 2010, it was discovered that the Pitted Ware possessed a very low level (5%) of an allele (−13910*T) strongly associated with the ability to consume unprocessed milk. This frequency is dramatically different from modern Swedes (74%). Whether the increase of this allele among the Swedes was a result of admixture or natural selection was uncertain. In a genetic study published in Science in April 2012, an individual from the Pitted Ware culture was examined. The individual was found to have “a genetic profile that is not fully represented by any sampled contemporary population”. In another genetic study published in Science in May 2014, the mtDNA of six individuals ascribed to the Pitted Ware culture was extracted. Four samples belonged to U4d, one belonged to U, and one belonged to V.” ref
“A genetic study published in August 2014 found that Pitted Ware peoples were closely genetically similar to people of the Catacomb culture, who like the Pitted Ware people carried high frequencies of the maternal haplogroups U5 and U4. These lineages are associated with Western Hunter-Gatherers and Eastern Hunter-Gatherers. In a genetic study published in Nature in September 2014, members of the Pitted Ware culture were determined to largely belong to the Scandinavian Hunter-Gatherer (SHG) cluster. In a genetic study published in Proceedings of the Royal Society B in January 2015, the mtDNA of thirteen PCW individuals from Öland and Gotland was extracted. The four individuals from Öland carried H1f, T2b, K1a1 and U4a1. Of the ten individuals from Gotland, four carried U4, two carried U5 haplotypes, two carried K1a1, and one carried HV0. The results indicated that the Pitted Ware culture was genetically distinct from the Funnelbeaker culture, and closely genetically related to earlier Mesolithic hunter-gatherers of Scandinavia and Western Europe. It was found that the Pitted Ware culture left a genetic imprint on Scandinavians, although this number is certainly not more than 60%.” ref
“A genetic study published in Nature Communications in January 2018 indicated genetic continuity between SHGs and the Pitted Ware culture, and found that the Pitted Ware people were genetically distinct from the Funnelbeaker culture. A 2018 study described the skin, hair and eye pigmentation of two Pitted Ware individuals from the Ajvide Settlement on the western coast of Gotland. The authors described the Pitted Ware specimens as sharing part of their phenotypic variation with Scandinavian Hunter Gatherers. Both specimens had a high probability of being dark-haired. One exhibited alleles associated with dark skin pigmentation on the SLC24A5 and SLC45A2 genes, while the other specimen carried a mix of alleles at these positions. The two specimens had a high probability of being blue eyed. A 2019 study published in Proceedings of the Royal Society B the remains of a Pitted Ware male were analyzed. He was found to the carrying the maternal haplgroup U5b1d2, and probably a subclade of the paternal haplogroup I2. He was estimated to be 25–35 years old and 165–175 cm tall. It was found that the Pitted Ware people only slightly contributed to the gene pool of the Battle Axe culture, who were almost wholly of Western Steppe Herder descent.” ref
“A genetic study published in Proceedings of the Royal Society B in June 2020 examined the remains of 19 Pitted Ware individuals buried on the island of Gotland. The study included a number of individuals who had been buried in a way typical of the Battle Axe culture. The 6 samples of Y-DNA extracted belonged to the paternal haplogroup I2a-L460 (2 samples), I2-M438 (2 samples), I2a1a-CTS595 and I2a1b1-L161. The 17 samples of mtDNA extracted belonged overwhelmingly to the maternal haplogroups U4 and U5. The study found no evidence of Battle Axe admixture among the Pitted Ware. They were genetically very different from earlier Funnelbeaker inhabitants of Gotland, although they carried a tiny amount of Early European Farmers (EEF) admixture. The evidence suggested that while the Pitted Ware culture was culturally influenced by the Battle Axe culture, it was not genetically influenced by it.” ref


Comb Ceramic culture
“The Comb Ceramic culture or Pit-Comb Ware culture, often abbreviated as CCC or PCW, was a northeast European culture characterised by its Pit–Comb Ware. It existed from around 4200 BCE to around 2000 BCE. The bearers of the Comb Ceramic culture are thought to have still mostly followed the Mesolithic hunter-gatherer (Eastern Hunter-Gatherer) lifestyle, with traces of early agriculture. The distribution of the artifacts found includes Finnmark (Norway) in the north, the Kalix River (Sweden) and the Gulf of Bothnia (Finland) in the west and the Vistula River (Poland) in the south. It would include the Narva culture of Estonia and the Sperrings culture in Finland, among others. They are thought to have been essentially hunter-gatherers, though e.g. the Narva culture in Estonia shows some evidence of agriculture. Some of this region was absorbed by the later Corded Ware horizon. The Pit–Comb Ware culture is one of the few exceptions to the rule that pottery and farming coexist in Europe. In the Near East farming appeared before pottery, then when farming spread into Europe from the Near East, pottery-making came with it. However, in Asia, where the oldest pottery has been found, pottery was made long before farming. It appears that the Comb Ceramic Culture reflects influences from Siberia and distant China.” ref
“By dating according to the elevation of land, the ceramics have traditionally (Äyräpää 1930) been divided into the following periods: early (Ka I, c. 4200 BC – 3300 BC), typical (Ka II, c. 3300 BC – 2700 BC) and late Comb Ceramic (Ka III, c. 2800 BC – 2000 BC). However, calibrated radiocarbon dates for the comb-ware fragments found (e.g., in the Karelian isthmus), give a total interval of 5600 BC – 2300 BC (Geochronometria Vol. 23, pp 93–99, 2004). The settlements were located at sea shores or beside lakes and the economy was based on hunting, fishing, and the gathering of plants. In Finland, it was a maritime culture that became more and more specialized in hunting seals. The dominant dwelling was probably a teepee of about 30 square meters where some 15 people could live. Also, rectangular houses made of timber became popular in Finland from 4000 BC cal. Graves were dug at the settlements and the dead were covered with red ochre. The typical Comb Ceramic age shows an extensive use of objects made of flint and amber as grave offerings.” ref
“The stone tools changed very little over time. They were made of local materials such as slate and quartz. Finds suggest a fairly extensive exchange network: red slate originating from northern Scandinavia, asbestos from Lake Saimaa, green slate from Lake Onega, amber from the southern shores of the Baltic Sea, and flint from the Valdai area in northwestern Russia. The culture was characterized by small figurines of burnt clay and animal heads made of stone. The animal heads usually depict moose and bears and were derived from the art of the Mesolithic. There were also many rock paintings. There are sources noting that the typical comb ceramic pottery had a sense of luxury and that its makers knew how to wear precious amber pendants. The great westward dispersal of the Uralic languages is suggested to have happened long after the demise of the Comb Ceramic culture, perhaps in the 1st millennium BC.” ref
“Saag et al. (2017) analyzed three CCC individuals buried at Kudruküla as belonging to Y-hg R1a5-YP1272 (R1a1b~ after ISOGG 2020), along with three mtDNA samples of mt-hg U5b1d1, U4a and U2e1. Mittnik (2018) analyzed two CCC individuals. The male carried R1 (2021: R1b-M343) and U4d2, while the female carried U5a1d2b. Generally, the CCC individuals were mostly of Eastern Hunter-Gatherer (EHG) descent, with even more EHG than people of the Narva culture. Lamnidis et al. (2018) found 15% Western Hunter-Gatherer (WHG) ancestry, 65% Eastern Hunter-Gatherer (EHG) – higher than among earlier cultures of the eastern Baltic, and 20% Western Steppe Herder (WSH).” ref


Postglacial genomes from foragers across Northern Eurasia reveal prehistoric
mobility associated with the spread of the Uralic and Yeniseian languages
Abstract
“The North Eurasian forest and forest-steppe zones have sustained millennia of sociocultural connections among northern peoples. We present genome-wide ancient DNA data for 181 individuals from this region spanning the Mesolithic, Neolithic, and Bronze Age. We find that Early to Mid-Holocene hunter-gatherer populations from across the southern forest and forest-steppes of Northern Eurasia can be characterized by a continuous gradient of ancestry that remained stable for millennia, ranging from fully West Eurasian in the Baltic region to fully East Asian in the Transbaikal region. In contrast, cotemporaneous groups in far Northeast Siberia were genetically distinct, retaining high levels of continuity from a population that was the primary source of ancestry for Native Americans. By the mid-Holocene, admixture between this early Northeastern Siberian population and groups from Inland East Asia and the Amur River Basin produced two distinctive populations in eastern Siberia that played an important role in the genetic formation of later people. Ancestry from the first population, Cis-Baikal Late Neolithic-Bronze Age (Cisbaikal_LNBA), is found substantially only among Yeniseian-speaking groups and those known to have admixed with them. Ancestry from the second, Yakutian Late Neolithic-Bronze Age (Yakutia_LNBA), is strongly associated with present-day Uralic speakers. We show how Yakutia_LNBA ancestry spread from an east Siberian origin ~4.5kya, along with subclades of Y-chromosome haplogroup N occurring at high frequencies among present-day Uralic speakers, into Western and Central Siberia in communities associated with Seima-Turbino metallurgy: a suite of advanced bronze casting techniques that spread explosively across an enormous region of Northern Eurasia ~4.0kya. However, the ancestry of the 16 Seima-Turbino-period individuals–the first reported from sites with this metallurgy–was otherwise extraordinarily diverse, with partial descent from Indo-Iranian-speaking pastoralists and multiple hunter-gatherer populations from widely separated regions of Eurasia. Our results provide support for theories suggesting that early Uralic speakers at the beginning of their westward dispersal where involved in the expansion of Seima-Turbino metallurgical traditions, and suggests that both cultural transmission and migration were important in the spread of Seima-Turbino material culture.” ref

Sidelkino Ancestry?
“The Sidelkino people in the east had genes associated with dark eyes and light skin.” ref
“The marked shift from Western Hunter-Gatherer-like towards Eastern Hunter-Gatherer-related ancestry from Baltic Mesolithic (ca. 30%) to Combed Ware cultures (ca. 65%-100%) supports this continuous westward expansion, that is possibly best represented in the currently available sampling by the ‘south-eastern’ shift (CHG:ANE-related) of the hunter-gatherer from Lebyazhinka IV (5600 BCE) relative to the older one from Sidelkino (9300 BCE), both from the Samara region in the Middle Volga.” ref
Dzudzuana, Sidelkino, and the Caucasus contribution to the Pontic-Caspian steppe
“It has been known for a long time that the Caucasus must have hosted many (at least partially) isolated populations, probably helped by geographical boundaries, setting it apart from open Eurasian areas. David Reich writes in his book the following about India: The genetic data told a clear story. Around a third of Indian groups experienced population bottlenecks as strong or stronger than the ones that occurred among Finns or Ashkenazi Jews. We later confirmed this finding in an even larger dataset that we collected working with Thangaraj: genetic data from more than 250 jati groups spread throughout India (…)” ref
Genetic ancestry changes in Stone to Bronze Age transition in the East European plain
“The transition from Stone to Bronze Age in Central and Western Europe was a period of major population movements originating from the Ponto-Caspian Steppe. Here, we report new genome-wide sequence data from 30 individuals north of this area, from the understudied western part of present-day Russia, including 3 Stone Age hunter-gatherers (10,800 to 4250 cal BCE) and 26 Bronze Age farmers from the Corded Ware complex Fatyanovo Culture (2900 to 2050 cal BCE). We show that Eastern hunter-gatherer ancestry was present in northwestern Russia already from around 10,000 BCE. Furthermore, we see a change in ancestry with the arrival of farming—Fatyanovo Culture individuals were genetically similar to other Corded Ware cultures, carrying a mixture of Steppe and European early farmer ancestry. Thus, they likely originate from a fast migration toward the northeast from somewhere near modern-day Ukraine—the closest area where these ancestries coexisted from around 3000 BCE.” ref
“The researchers studied the genetic affinities of the higher-coverage (5×) HG PES001 further using outgroup f3 statistics by comparing the affinities of three higher-coverage Russian Mesolithic HGs—PES001 (Peschanitsa; 10,785 to 10,626 cal BCE), I0061 (Yuzhnyy Oleni Ostrov; 6773 to 5886 BCE), and Sidelkino (Sidelkino; 9386 to 9231 cal BCE)—to Mesolithic and Paleolithic HGs from different areas of Europe and Siberia (Fig. 2C and table S9). All three are most similar to individuals from the European part of Russia or from Siberia who lived within 10,000 years of each other—to each other, the West Siberia Neolithic population, and the Afontova Gora 3 individual. These are followed by individuals from Central Europe from the same time window. Geographically close Paleolithic Sunghir and Kostenki individuals from >30,000 cal BCE share less than temporally close HGs from Central Europe with the Russian Mesolithic individuals. We also tried modeling PES001 as a mixture of WHG and either CHG, Mal’ta, or Afontova Gora 3 using qpAdm, but all three models were rejected (P value of 1.96 × 10−85 to 7.93 × 10−13).” ref
EF ancestry in Fatyanovo Culture individuals
“Then, the researchers turned to the Bronze Age Fatyanovo Culture individuals and determined that they carry maternal (subclades of mtDNA hg U5, U4, U2e, H, T, W, J, K, I, and N1a) and paternal (chrY hg R1a-M417) lineages that have also been found in CWC individuals elsewhere in Europe. In all individuals for which the chrY hg could be determined with sufficient depth (n = 6), it is R1a2-Z93, a lineage now spread in Central and South Asia, rather than the R1a1-Z283 lineage that is common in Europe. R1a2-Z93 is also not rejected for the individuals that were determined with less depth due to missing data on more apical markers.” ref
“On the PCA, the Fatyanovo individuals (and the Estonian CWC individual) group together with many European Late Neolithic/Bronze Age (LNBA) and Steppe Middle/Late Bronze Age individuals on top of modern Northern and Eastern Europeans (Fig. 2A). This ancient cluster is shifted toward Anatolian and European EF compared to Steppe Early/Middle Bronze Age populations, including the Yamnaya. The same could be seen in ADMIXTURE analysis where the Fatyanovo individuals are most similar to LNBA Steppe ancestry populations from Central Europe, Scandinavia, and the Eastern Baltic (Fig. 2B). These populations are composed of the “WHG” (blue) and “ancient Caucasus/Iran” (olive) component and small amounts of the “Russian Far East” (orange) component, similarly to Yamnaya populations. However, the European LNBA populations (including Fatyanovo) also display a component most frequent in Anatolian and European EF populations (light green), which is not present in the Yamnaya from Russia.” ref
“The researchers studied the affinities of the Fatyanovo individuals by comparing FST, outgroup f3, and D statistics’ results of different populations and found that Fatyanovo shares more with European EF populations and modern Near Easterners than Yamnaya Samara does (Fig. 3C). This signal can also be seen when using either autosomal or X chromosome (chrX) positions from the 1240K dataset instead of the autosomal positions of the HO dataset with less single-nucleotide polymorphisms (SNPs). We also compared the affinities of Yamnaya Samara and Fatyanovo directly with D statistics and saw that Fatyanovo is significantly more similar (Z > 3) to most EF populations than to Yamnaya Samara, and the latter, in turn, is significantly more similar to most Steppe populations than to Fatyanovo. We studied the apparent EF input in Fatyanovo further using admixture f3 statistics and got significant results (Z < −3) for admixture between different Yamnaya groups and a wide variety of EF populations. Furthermore, when comparing outgroup f3 and D statistics’ results for Fatyanovo and Central CWC, there are no clear differences in their affinities to different ancient or modern population groups (Fig. 3D).” ref
“Because the previous analyses suggested that the genetic makeup of the Fatyanovo Culture individuals was a result of admixture between migrating Yamnaya individuals and contemporary European populations, we used two complementary methods (qpAdm and ChromoPainter/NNLS) to determine suitable proxies for the admixing populations and the mixing proportions (Fig. 4). We tested qpAdm models including Yamnaya from Samara or Kalmykia and a variety of EF populations one at a time and found that the two EF populations, with the highest P values with both Yamnaya populations, are Globular Amphora and Trypillia (P values of 0.02/0.16 and 0.06/0.26, respectively) (table S15). The admixture proportions are 65.5%/66.9% Yamnaya Samara/Kalmykia + 34.5%/33.1% Globular Amphora and 65.5%/69.6% Yamnaya Samara/Kalmykia + 34.5%/30.4% Trypillia, respectively (Fig. 4). The proportions are similar (69 to 75% Yamnaya + 25 to 31% EF) for Central and Baltic CWC (Fig. 4).” ref
“We also tested these four models with the preceding Volosovo Culture HG BER001 added to “right” populations to see whether there is shared drift between Volosovo and Fatyanovo that would cause the models with admixing populations without this drift to be rejected. All four models are still not rejected (P values of 0.97/0.57 and 0.98/0.59, respectively), suggesting that no Volosovo contribution is needed to model Fatyanovo. Ancestry proportions were also calculated using the ChromoPainter/NNLS pipeline with the results 37%/38% Yamnaya Samara + 63%/62% Globular Amphora/Trypillia for Fatyanovo and 51 to 60% Yamnaya + 40 to 49% EF for Central/Baltic CWC.” ref
“Although the estimated proportion of EF ancestry is higher for Fatyanovo compared to the other groups in both cases, the difference is significant (P value of 0.005 to 0.03) only with Trypillia in the model. Note that qpAdm calculates admixture between populations, while ChromoPainter/NNLS uses single individuals as sources, which might influence the results. Although two-way admixture between Yamnaya and an EF population is enough to explain the genetic variation in Fatyanovo, qpAdm models with an HG population added are also not rejected with EHG, WeRuHG, and Volosovo (P values of 0.14 to 0.98). Fatyanovo can be modeled as 60 to 63% Yamnaya Samara + 33 to 34% Globular Amphora + 3 to 6% HG. The results are similar for Central and Baltic CWC with 2 to 11% of HG ancestry (except with Volosovo as a source for Central CWC, which gets a negative mixture coefficient).” ref
“From around 8,00 years ago, we begin to observe admixture events with Sidelkino ancestry in central Europe. This is first detected in an individual from Gross Fredenwalde in northeastern Germany and reaches around 10% in most European hunter-gatherer individuals thereafter. Soon after 8 ka, Sidelkino ancestry was absent in eastern Spain but it had already reached northern Iberia alongside an increase in Oberkassel ancestry (Fig. 5). Conversely, additional Oberkassel ancestry is identified in eastern Europe by at least 7,500 years ago in newly generated genomes from Minino I and Yazykovo from the upper Volga region, whereas a 1,000-years-older individual from Minino I did not have this genetic component. Considering a freshwater reservoir signal in the upper Volga region making radiocarbon dates on human remains appear up to about 500 years older than their true age, there could be an interval of more than 1,000 years between the first evidence of admixture in central European hunter-gatherers with Sidelkino ancestry and eastern European hunter-gatherers with Oberkassel ancestry. However, additional genomes intermediate in time and space are needed to assess whether those two admixture events were independent or part of a common demographic process.” ref
“After 7,500 years ago, as ANF ancestry had reached regions north of the Alps, individuals carrying a hunter-gatherer genetic profile were primarily restricted to the northern fringes of Europe (Fig. 5). In this period, the Oberkassel-ancestry admixture spread further east, reaching Samara by around 6,500 years ago, and an increase in Sidelkino ancestry was detected in hunter-gatherers from the Baltic region, which was previously associated with the transition from the Narva culture to the Comb Ceramic culture (Fig. 8). In central Europe, admixture with ANF ancestry became highly common but not ubiquitous, indicating the co-existence of hunter-gatherer and farmer societies without admixing for several hundred years. The youngest individual carrying large portions of hunter-gatherer ancestry in the analyzed dataset is from Ostorf in northern Germany, dated to around 5,200 years ago (>90% Oberkassel cluster plus Sidelkino-cluster components). Individuals at this site might mark one of the last occurrences of such high levels of hunter-gatherer-related ancestries, just centuries before the emerging European Bronze Age.” ref
“On the basis of PCA and outgroup f3-statistics, the Neolithic Tutkaul 1 individual from Tajikistan is closely related to Upper Palaeolithic individuals from south-central Siberia (Afontova Gora 3 (AG3) and Mal’ta 1), and roughly contemporaneous West Siberian hunter-gatherers (Tyumen and Sosnoviy), both carrying high proportions of ANE ancestry (Fig. 1c and Fig. 6). We tested the affinity of Tutkaul 1 to worldwide ancient and modern populations relative to AG3. Contrary to West Siberian hunter-gatherers, Tutkaul 1 does not carry an extra eastern Eurasian ancestry, but shows affinity to Iranian Neolithic farmers and some younger populations from Iran and the Turan region. Conversely, individuals in the Sidelkino cluster are genetically closer to AG3 than Tutkaul 1. This suggests that the newly reported Neolithic individual from central Asia carries an ancestry that might be a good proxy for the ANE-related contribution to Iran and the Turan region from around 5,500 years ago but not to roughly contemporaneous hunter-gatherers from eastern Europe.” ref
“In sum, we describe the formation and interaction between the Oberkassel and Sidelkino clusters, the two main hunter-gatherer ancestries present in Europe from 14 ka onwards. The genomic similarity of the Oberkassel cluster to Arene Candide 16 in northwestern Italy might imply that Epigravettian-associated ancestry spread from the south to central Europe passing through the western side of the Alpine region. The Sidelkino ancestry also emerged around 14,000 years ago with its first direct evidence in eastern Europe dated to 13,000 years ago. The increasing level of admixture between distinct hunter-gatherer populations from around 8,000 years ago onwards indicates an intensified mobility of those forager groups. This might have been in part triggered by the concomitant expansion of Neolithic farmers across Europe and/or by environmental factors, such as the climatic event around 8,200 years ago, the largest abrupt cooling in the northern hemisphere during the Holocene epoch.” ref
“Leveraging the substantially increased sample size, we investigated genetically distinct hunter-gatherer groups for allele frequencies at selected loci that are known to be associated with specific phenotypic traits in present-day Europeans (Fig. 5b, Figs. 27 and 28). Consistent with previous findings, none of the analyzed groups show the derived allele at SNP rs4988235 on the LCT gene, which is responsible for lactase persistence. As previously hinted, we find a large frequency variation in alleles related to skin and eye pigmentation among post-LGM hunter-gatherer groups. For the SNP associated with light eye color (HERC2/OCA2 (rs12913832)), individuals from the Villabruna cluster, Oberkassel cluster, Baltic HG, and SHG groups show high frequencies of the derived allele (>90%), which is responsible for the green or blue eye phenotype, whereas Sidelkino cluster, Ukraine HG and Iron Gates HG groups show the low occurrence of this allele (10–25%).” ref
“Instead, for the two SNPs associated with skin color (SLC24A5 (rs1426654) and SLC45A2 (rs16891982)), Sidelkino cluster and Ukraine HG groups show a higher frequency (>90% for SLC24A5 and 29–61% for SLC45A2) of the derived alleles related to light skin color, compared with Oberkassel and Villabruna clusters, where those alleles are almost completely absent (<1%). On the basis of the genetic variation of present-day Europeans, this could imply phenotypic differences between post-14 ka hunter-gatherer populations across Europe, with individuals in the Oberkassel cluster possibly exhibiting darker skin and lighter eyes, and individuals in the Sidelkino cluster possibly lighter skin and darker eye color.” ref
The data generated in this study enabled us to investigate genomic transformations of and interactions between Eurasian hunter-gatherers at high resolution (Fig. 9). We provide five novel insights into the genomic history of hunter-gatherer populations over a time span of 30,000 years from the Upper Palaeolithic to the Neolithic. First, we show that individuals associated with the Gravettian culture across Europe were not a biologically homogeneous population. Culturally, however, we see both widespread general tendencies, such as weaponry and some portable art, and other aspects that have a more regional character, such as mortuary practices, various originalities in lithic and hard organic materials tool kits and adornments.” ref
“The ancestry found in individuals associated with the preceding Aurignacian culture from central Europe (GoyetQ116-1 ancestry) gave rise to Gravettian-associated individuals from western and southwestern Europe. This derived ancestry—the Fournol cluster—survived during the LGM in Solutrean-associated individuals, possibly within the Franco-Cantabrian climatic refugium, leading to later populations associated with the Magdalenian culture (GoyetQ2 cluster and El Mirón). Conversely, the ancestry found in pre-30 ka eastern European individuals (Kostenki cluster and Sunghir group) contributed to Gravettian-associated individuals from central and southern Europe (Věstonice cluster), the latter without descendants retrieved in post-LGM populations from those regions.” ref
“Second, the ancestry of individuals associated with the Epigravettian culture (Villabruna cluster), which was found to genetically connect European and Near Eastern hunter-gatherers, reached southern Europe well before the transition between the Early and Late Epigravettian and possibly as early as the Gravettian–Epigravettian transition. A phylogeographic reconstruction of different lineages carrying this ancestry further suggests its entry point into northeastern Italy from the Balkans followed by a north-to-south expansion into the Italian peninsula alongside a population decline through sequential bottlenecks.” ref
“Third, Magdalenian-associated individuals not only from Iberia but also from the rest of Europe carry Epigravettian-associated ancestry (Villabruna cluster). Genetic analyses of western European individuals associated with the preceding Badegoulian culture might provide clues on the processes that led to the formation of the GoyetQ2 cluster. As inferred from the archaeological record, the spread of the Magdalenian across Europe is linked to southwestern to northern and northeastern post-LGM population expansions and not to movements from southeastern refugia.” ref
“Fourth, we extend the finding of a large-scale genetic turnover as early as 14 ka in central and western European hunter-gatherers associated with multiple techno-complexes—Federmesser, Azilian, and other Final Palaeolithic groups—despite considerable technological continuity with the preceding late Magdalenian. This broadly distributed ancestry (the Oberkassel cluster (also known as WHG)) is most closely related to an Epigravettian-associated individual from northwestern Italy, suggesting that its expansion into continental Europe might have started from the west—and not the east—side of the Alps. Moreover, the almost complete genetic replacement of the Magdalenian-associated gene pool raises the hypothesis that parts of Europe were differentially populated during the abrupt climatic variation starting around 14,700 years ago with the Bølling–Allerød warming period, creating areas where southern European populations could expand. This might also explain the genetic uniformity of the Oberkassel cluster across large parts of western Eurasia but genomic data from between 15,000 and 14,000 years ago is needed to understand the exact dynamics of this turnover.” ref
“Fifth, the Oberkassel ancestry in western and central Europe and the Sidelkino ancestry in eastern Europe remained largely isolated for almost 6,000 years until genetic interactions were first observed—around 8,000 years ago in northeastern Germany, possibly associated with cultural exchanges along the Baltics and around 7,5000 years ago in the upper Volga region, possibly linked to the spread of pottery in the region. This study reveals that western and southwestern Europe served as climatic refugia for the persistence of human groups during the coldest phase of the last Ice Age whereas populations in the Italian peninsula and the eastern European plain were genetically overturned, challenging the role of these regions as glacial refugia for humans.” ref
“The incoming Villabruna ancestry later became the most widespread hunter-gatherer ancestry across Europe. Further palaeogenomic studies on Upper Palaeolithic individuals from the Balkans will be essential for understanding whether southeastern Europe represents the source of the Villabruna ancestry and a climatic refugium for human populations during the LGM. Note added in proof: A companion paper describes genome-wide data of a 23,000-year-old Solutrean-associated individual from southern Iberia that extend the evidence of genetic continuity across the LGM in southwestern Europe.” ref
“The Yamnaya archaeological complex appeared around 3300BCE across the steppes north of the Black and Caspian Seas, and by 3000BCE reached its maximal extent from Hungary in the west to Kazakhstan in the east. To localize the ancestral and geographical origins of the Yamnaya among the diverse Eneolithic people that preceded them, we studied ancient DNA data from 428 individuals of which 299 are reported for the first time, demonstrating three previously unknown Eneolithic genetic clines. First, a “Caucasus-Lower Volga” (CLV) Cline suffused with Caucasus hunter-gatherer (CHG) ancestry extended between a Caucasus Neolithic southern end in Neolithic Armenia, and a steppe northern end in Berezhnovka in the Lower Volga. Bidirectional gene flow across the CLV cline created admixed intermediate populations in both the north Caucasus, such as the Maikop people, and on the steppe, such as those at the site of Remontnoye north of the Manych depression. CLV people also helped form two major riverine clines by admixing with distinct groups of European hunter-gatherers.” ref
“A “Volga Cline” was formed as Lower Volga people mixed with upriver populations that had more Eastern hunter-gatherer (EHG) ancestry, creating genetically hyper-variable populations as at Khvalynsk in the Middle Volga. A “Dnipro Cline” was formed as CLV people bearing both Caucasus Neolithic and Lower Volga ancestry moved west and acquired Ukraine Neolithic hunter-gatherer (UNHG) ancestry to establish the population of the Serednii Stih culture from which the direct ancestors of the Yamnaya themselves were formed around 4000BCE. This population grew rapidly after 3750–3350BCE, precipitating the expansion of people of the Yamnaya culture who totally displaced previous groups on the Volga and further east, while admixing with more sedentary groups in the west. CLV cline people with Lower Volga ancestry contributed four fifths of the ancestry of the Yamnaya, but also, entering Anatolia from the east, contributed at least a tenth of the ancestry of Bronze Age Central Anatolians, where the Hittite language, related to the Indo-European languages spread by the Yamnaya, was spoken. We thus propose that the final unity of the speakers of the “Proto-Indo-Anatolian” ancestral language of both Anatolian and Indo-European languages can be traced to CLV cline people sometime between 4400–4000 BCE.” ref
“The origin of Indo-Anatolian and Indo-European languages. Genetic reconstruction of the ancestry of Pontic-Caspian steppe and West Asian populations points to the North Caucasus-Lower Volga area as the homeland of Indo-Anatolian languages and to the Serednii Stih archaeological culture of the Dnipro-Don area as the homeland of Indo-European languages. The Caucasus-Lower Volga people had diverse distal roots, estimated using the qpAdm software on the left barplot, as Caucasus hunter-gatherer (purple), Central Asian (red), Eastern hunter-gatherer (pink), and West Asian Neolithic (green). Caucasus-Lower Volga expansions, estimated using qpAdm on the right barplot as disseminated Caucasus Neolithic (blue)-Lower Volga Eneolithic (orange) proximal ancestries, mixing with the inhabitants of the North Pontic region (yellow), Volga region (yellow), and West Asia (green).” ref
“Between 3300–1500 BCE, people of the Yamnaya archaeological complex and their descendants, in subsequent waves of migration, spread over large parts of Eurasia, contributing to the ancestry of people of Europe, Central and South Asia, Siberia, and the Caucasus. The spread of Indo-European language and culture transformed all these regions. Despite the centrality of the Yamnaya expansion to the human story of Bronze Age Eurasia, their ancestral origins are poorly understood. A first challenge has been the sparse sampling of the Yamnaya themselves across their enormous geographic distribution. The remarkable long-range mobility of the Yamnaya, quickly spreading over a vast region, adds further difficulties to tracing, from radiocarbon dating, the origins of their material culture and associated genetic profile. Nor can these origins be traced to the numerous earlier Eneolithic cultures that preceded the Yamnaya, and among whom their ancestors must be sought, as these have been sampled even more poorly and systematically.” ref
“The first formal study of the origins of the Yamnaya identified two disparate sources of ancestry: a northern, “Eastern Hunter-Gatherer” (EHG) source from far eastern Europe, and a southern, West Asian source related to present-day Armenians.The latter source was revealed, by ancient DNA, to be related to some of the region’s earliest inhabitants: Paleolithic-Mesolithic “Caucasus Hunter-Gatherers” (CHG) of Georgia, and Neolithic people of the Zagros and South Caucasus. Additional discoveries further complicated the stories of both the northern and southern ancestors of the Yamnaya. First, it was noted that both CHG and EHG were part of an interaction sphere across the boundary between West Asia and eastern Europe, suggesting the existence of intermediate populations and raising the question of when and where these came together to form the Eneolithic antecessors of the Yamnaya. Second, it was recognized that the steppe itself was an admixture zone of EHG with “Western Hunter-Gatherers” (WHG). Mesolithic hunter-gatherers from Ukraine were succeeded by more WHG-admixed Neolithic hunter-gatherers in the Dnipro valley, representing a local reshuffling within the European portion of a ~7,000 km-long trans-Eurasian cline of boreal hunter-gatherers.” ref
“What was the relative contribution of the EHG (who were present in the Volga River at Lebyazhinka ca. 5660–5535 BCE) and these more western Ukraine Neolithic hunter-gatherers (UNHG) of the Dnipro to later populations? Third, it was discovered that the Yamnaya had not only CHG-related, but also Anatolian Neolithic ancestry, absent in the early known steppe inhabitants, and derived from European farmer neighbors west of the steppe. This ancestry was later shown to be of rather Anatolian-Levantine-Mesopotamian origin, and to be mediated not from Europe but from the Caucasus neighbors south of the steppe. Such ancestry must have been added following the expansion of Neolithic farmers into the Caucasus, introduced thence into the steppe as a later exogenous element, distinct from the earlier CHG-related one. Finally, it was recognized that European steppe populations were formed not only by northern-southern admixture, but included, in at least some Eneolithic and Bronze Age people of the North Caucasus, contributions related to Siberians from further east. What was the extent of the spread of this eastern ancestry, and did the Yamnaya themselves possess it?” ref
“Here we present a unified population genetic analysis of 372 newly reported individuals dating from 6400–2000 BCE, as well as increased quality data for 61 individuals. The present study serves as the formal technical report for 299 of the newly reported individuals and 55 of the individuals with increased quality data; more than 80% of the individuals are from Russia, but the dataset is also significant in including dozens of individuals from westward expansion of Steppe cultures along the Danube.” ref
“A parallel study presents a combined archaeological and genetic analysis of population transformations in the North Pontic Region (Ukraine and Moldova) and serves as the formal report for the data from the other 73 of the newly analyzed individuals and the other 5 individuals with increased quality data, with both studies co-analyzing the full dataset. We grouped individuals into analysis labels based on geographical and temporal information, archaeological context, and genetic clustering. The potential of the combined dataset for shedding light on this period can be appreciated from the fact that it adds 79 analyzed Eneolithic people from the steppe and its environs (from Russia or Ukraine, west of 60E longitude and south of 60N latitude, between 5000–3500BCE) to 82 published and a total of 286 Yamnaya/Afanasievo individuals compared to 75 in the literature.” ref
J-M172 (Zagros mountains origin)
“J-M304 (Transcaucasian origin) is defined by the M304 genetic marker, or the equivalent 12f2.1 marker. The main current subgroups J-M267 (Armenian highlands origin) and J-M172 (Zagros mountains origin), which now comprise between them almost all of the haplogroup’s descendant lineages, are both believed to have arisen very early, at least 10,000 years ago. Nonetheless, Y-chromosomes F-M89* and IJ-M429* were reported to have been observed in the Iranian plateau (Grugni et al. 2012). On the other hand, it would seem to be that different episodes of populace movement had impacted southeast Europe, as well as the role of the Balkans as a long-standing corridor to Europe from the Near East is shown by the phylogenetic unification of Hgs I and J by the basal M429 mutation. This proof of common ancestry suggests that ancestral Hgs IJ-M429* probably would have entered Europe through the Balkan track sometime before the LGM. They then subsequently split into Hg J and Hg I in Middle East and Europe in a typical disjunctive phylogeographic pattern.” ref
“Haplogroup J-M172 (J2-M172) is common in modern populations in Western Asia, Central Asia, South Asia, Southern Europe, Northwestern Iran and North Africa. It is thought that J-M172 may have originated in the Caucasus, Anatolia and/or Western Iran. The date of origin for haplogroup J-M172 was estimated by Batini et al in 2015 as between 19,000 and 24,000 years ago. Samino et al in 2004 dated the origin of the parent haplogroup, J-P209, to between 18,900 and 44,500 years ago. Ancient J-M410, specifically subclade J-Y12379*, has been found, in a mesolithic context, in a tooth from the Kotias Klde Cave in western Georgia dating 9.529-9.895 years ago. This sample has been assigned to the Caucasus hunter-gatherers (CHG) autosomal component. J-M410, more specifically its subclade J-PF5008, has also been found in a mesolithic sample from the Hotu and Kamarband Caves located in Mazandaran Province of Iran, dating back to 9,100-8,600 BCE (approximately 11,000 yrats ago). Both samples belong to the Trialetian Culture. It is likely that J2 men had settled over most of Anatolia, the South Caucasus and the Zagros mountains by the end of the Last Glaciation 12,000 years ago.” ref
“Zalloua & Wells (2004) and Al-Zahery et al. (2003) claimed to have uncovered the earliest known migration of J2, expanded possibly from Anatolia and the Caucasus. Nebel et al. (2001) found that, “According to Underhill et al. (2000), Eu 9 (H58) evolved from Eu 10 (H71) through a T→G transversion at M172 (emphasis added),” and that in today’s populations, Eu 9 (the post-mutation form of M172) is strongest in the Caucasus, Asia Minor and the Levant, whilst Eu 10 becomes stronger and replaces the frequency of Eu 9 as one moves south into the Arabian Peninsula, so that people from the Caucasus met with Arabs near and between Mesopotamia (Sumer/Assyria) and the Negev Desert, as “Arabisation” spread from Arabia to the Fertile Crescent and Turkey.” ref
“Di Giacomo et al. (2004) postulated that J-M172 haplogroup spread into Southern Europe from either the Levant or Anatolia, likely parallel to the development of agriculture. As to the timing of its spread into Europe, Di Giacomo et al. points to events which post-date the Neolithic, in particular the demographic floruit associated with the rise of the Ancient Greek world. Semino et al. (2004) derived older age estimates for overall J2 (having used the Zhivotovsky method c.f. Di Giacomo), postulating its initial spread with Neolithic farmers from the Near East. However, its subclade distribution, showing localized peaks in the Southern Balkans, southern Italy, north/central Italy and the Caucasus, does not conform to a single ‘wave-of-advance’ scenario, betraying a number of still poorly understood post-Neolithic processes which created its current pattern. Like Di Giacomo et al., the Bronze Age southern Balkans was suggested by Semino et al. to have been an important vector of spread.” ref
“Haplogroup J-M172 is found mainly in the Fertile Crescent, the Caucasus, Anatolia, Italy, the Mediterranean littoral, and the Iranian plateau. Y-DNA: J2 (J-M172): Syrid/Nahrainid Arabid(s). J-M172 is found at very high frequencies in certain peoples of the Caucasus: among the Ingush 87.4%, Chechens 55.2%, Georgians 21%-72%, Azeris 24%-48%, Abkhaz 25%, Balkars 24%, Ossetians 24%, Armenians 21%-24%, Adyghe 21.8%, and other groups. J-M172 is typical of populations of the Near East, Southern Europe, Southwest Asia and the Caucasus, with a moderate distribution through much of Central Asia, South Asia, and North Africa. The highest reported frequency of J-M172 ever was 87.4%, among Ingush in Malgobek.” ref
“More specifically it is found in Iraq, Kuwait, Syria, Lebanon, Turkey, Georgia, Azerbaijan, North Caucasus, Armenia, Iran, Israel, Palestine, Cyprus, Greece, Albania, Italy, Spain, and more frequently in Iraqis 24%, Chechens 51.0%-58.0%, Georgians 21%-72%, Lebanese 30%, Ossetians 24%, Balkars 24%, Syrians 23%, Turks 13%-40%, Cypriots 12.9%-37%, Armenians 21%-24%, Circassians 21.8%, Iranians 10%-25%, Albanians 16%, Italians 9%-36%, Sephardi Jews 15%-29%, Maltese 21%, Palestinians 17%, Saudis 14%, Jordanians 14%, Omanis 10%-15%, and North Indian Shia Muslim 18%.” ref
“It has been proposed that haplogroup subclade J-M410 was linked to populations on ancient Crete by examining the relationship between Anatolian, Cretan, and Greek populations from around early Neolithic sites in Crete. Haplogroup J-M172 was associated with Neolithic Greece (ca. 8500 – 4300 BCE) and was reported to be found in modern Crete (3.1%) and mainland Greece (Macedonia 7.0%, Thessaly 8.8%, Argolis 1.8%). Sephardi Jews have about 15%-29%, of haplogroup J-M172, and Ashkenazi Jews have 15%-23%. It was reported in an early study which tested only four STR markers that a small sample of Italian Cohens belonged to Network 1.2, an early designation for the overall clade now known as J-L26, defined by the deletion at DYS413. However, a large number of all Jewish Cohens in the world belong to haplogroup J-M267 (see Cohen modal haplotype).” ref
“Haplogroup J-M172 has been shown to have a more northern distribution in the Middle East, although it exists in significant amounts in the southern middle-east regions, a lesser amount of it was found when compared to its brother haplogroup, J-M267, which has a high frequency southerly distribution. It was believed that the source population of J-M172 originated from the Levant/Syria (Syrid-J-M172), and that its occurrence among modern populations of Europe, Central Asia, and South Asia was a sign of the neolithic agriculturalists. However, as stated it is now believed more likely to have been spread in waves, as a result of post-Neolithic processes.” ref
“Haplogroup J-M172 is found in the highest concentrations in the Caucasus and the Fertile Crescent/Iraq and is found throughout the Mediterranean (including the Italian, Balkan, Anatolian and Iberian peninsulas and North Africa). The highest ever reported concentration of J-M172 was 72% in Northeastern Georgia. Other high reports include Ingush 32% (Nasidze 2004), Cypriots 30-37% (Capelli 2005), Lebanese 30% (Wells et al. 2001), Assyrian, Mandean and Arab Iraqis 29.7%, Syrians and Syriacs 22.5%, Kurds 24%-28%, Pashtuns 20-30%, Iranians 23% (Aburto 2006), Ashkenazi Jews 24%, Palestinian Arabs 16.8%-25%, Sephardic Jews 29% and North Indian Shia Muslim 18%, Chechens 26%, Balkars 24%, Yaghnobis 32%, Armenians 21-24%, and Azerbaijanis 24%-48%.” ref
“In South Asia, J2-M172 was found to be significantly higher among Dravidian castes at 19% than among Indo-European castes at 11%. J2-M172 and J-M410 is found 21% among Dravidian middle castes, followed by upper castes, 18.6%, and lower castes 14%. Subclades of M172 such as M67 and M92 were not found in either Indian or Pakistani samples which also might hint at a partial common origin.” ref
“According to a genetic study in China by Shou et al., J2-M172 is found with high frequency among Uygurs (17/50 = 34%) and Uzbeks (7/23 = 30.4%), moderate frequency among Pamiris (5/31 = 16.1%), and also found J-M172 in Han Chinese (10%) and low frequency among Yugurs (2/32 = 6.3%) and Monguors (1/50 = 2.0%). The authors also found J-M304(xJ2-M172) with low frequency among the Russians (1/19 = 5.3%), Uzbeks (1/23 = 4.3%), Sibe people (1/32 = 3.1%), Dongxiangs (1/35 = 2.9%), and Kazakhs (1/41 = 2.4%) in Northwest China. Only far northwestern ethnic minorities had haplogroup J in Xinjiang, China. Uzbeks in the sample had 30.4% J2-M172 and Tajiks of Xinjiang and Uyghurs also had it.” ref
“The oldest evidence of agriculture comes from the Fertile Crescent (~11,000 years ago), the centre for demic diffusion. However, evidences of first agriculture from South Asia indicate a timeline (~9,000-10,000 years ago) closer to the emergence of agriculture in the Fertile Crescent. Previous studies have established that J2a-M410 and J2b-M102 represent the Y-chromosomal component associated with demic diffusion of Neolithic farmers in North Africa and Eurasia from Mesopotamia (Iraq and Syria). The J2-M172 has been associated with different cultures and populations in history, such as Mediterranean/Aegean, Greco-Anatolian, Mesopotamian, and Caucasian. Presence of J2a-M410 and J2b-M102 in India has been considered a result of gene influx from Western Asia. Worldwide spatial distribution of haplogroup (HG) J2a-M410 coincides with presence of archaeological records of painted pottery and ceramic figurine culture. Similar material culture dating ~9,000 years ago has been recovered from the Neolithic sites of Mehrgarh located West of Indus Valley (now in Pakistan).” ref
“The population of the sub-continent shows an outstanding biological and cultural variation, which has been shaped by geographical, ecological, social, and linguistic factors. Many recent studies, particularly from the field of evolutionary genetics, have shown the extent to which each of these factors has contributed to the generation and maintenance of this diversity. It is pertinent that paternal gene pool of India comprises mainly HGs of autochthonous origin of Late Pleistocene ancestry and received very little gene flow from outside. Many Y chromosomal studies of India addressing major demographic questions have included and broadly discussed HG J2-M172.” ref
“However, the arrival and distribution of HG J2-M172 subclades in India have not been studied comprehensively yet. Hence, the aim of this study is to address the following: (1) whether or not, the demic diffusion model stands true for the current distribution of J2-M172 in India in the backdrop of independent origin of agriculture in South Asia; (2) which contemporary populations have the closest affinities with J2-M172 in India; (3) whether distribution of J2a-M410 in India is more dominant in castes than in tribes; and (4) what social, linguistic or geographical factors might have influenced the distribution of J2-M172 subclades. To address the above, we investigated the presence of different subclades of HG J2-M172 in diverse set of Indian populations and their affinities with rest of the world.” ref
Origin, Diffusion, and Differentiation of Y-Chromosome Haplogroups E and J: Inferences on the Neolithization of Europe and Later Migratory Events in the Mediterranean Area
“Abstract: The phylogeography of Y-chromosome haplogroups E (Hg E) and J (Hg J) was investigated in >2,400 subjects from 29 populations, mainly from Europe and the Mediterranean area but also from Africa and Asia. The observed 501 Hg E and 445 Hg J samples were subtyped using 36 binary markers and eight microsatellite loci. Spatial patterns reveal that (1) the two sister clades, J-M267 and J-M172, are distributed differentially within the Near East, North Africa, and Europe; (2) J-M267 was spread by two temporally distinct migratory episodes, the most recent one probably associated with the diffusion of Arab people; (3) E-M81 is typical of Berbers, and its presence in Iberia and Sicily is due to recent gene flow from North Africa; (4) J-M172(xM12) distribution is consistent with a Levantine/Anatolian dispersal route to southeastern Europe and may reflect the spread of Anatolian farmers; and (5) E-M78 (for which microsatellite data suggest an eastern African origin) and, to a lesser extent, J-M12(M102) lineages would trace the subsequent diffusion of people from the southern Balkans to the west. A 7%–22% contribution of Y chromosomes from Greece to southern Italy was estimated by admixture analysis.” ref



“The arrival of haplogroup R1a-M417 in Eastern Europe, and the east-west diffusion of pottery through North Eurasia.” ref
R-M417 (R1a1a1)
“R1a1a1 (R-M417) is the most widely found subclade, in two variations which are found respectively in Europe (R1a1a1b1 (R-Z282) ([R1a1a1a*] (R-Z282) and Central and South Asia (R1a1a1b2 (R-Z93) ([R1a1a2*] (R-Z93).” ref
R-Z282 (R1a1a1b1a) (Eastern Europe)
“This large subclade appears to encompass most of the R1a1a found in Europe.
- R1a1a1b1a [R1a1a1a*] (R-Z282*) occurs in northern Ukraine, Belarus, and Russia at a frequency of c. 20%.
- R1a1a1b1a3 [R1a1a1a1] (R-Z284) occurs in Northwest Europe and peaks at c. 20% in Norway.
- R1a1a1c (M64.2, M87, M204) is apparently rare: it was found in 1 of 117 males typed in southern Iran.” ref
R1a1a1b2 (R-Z93) (Asia)
“This large subclade appears to encompass most of the R1a1a found in Asia, being related to Indo-European migrations (including Scythians, Indo-Aryan migrations, and so on).
- R-Z93* or R1a1a1b2* (R1a1a2* in Underhill (2014)) is most common (>30%) in the South Siberian Altai region of Russia, cropping up in Kyrgyzstan (6%) and in all Iranian populations (1-8%).
- R-Z2125 occurs at highest frequencies in Kyrgyzstan and in Afghan Pashtuns (>40%). At a frequency of >10%, it is also observed in other Afghan ethnic groups and in some populations in the Caucasus and Iran.
- R-M434 is a subclade of Z2125. It was detected in 14 people (out of 3667 people tested), all in a restricted geographical range from Pakistan to Oman. This likely reflects a recent mutation event in Pakistan.
- R-M560 is very rare and was only observed in four samples: two Burushaski speakers (north Pakistan), one Hazara (Afghanistan), and one Iranian Azerbaijani.
- R-M780 occurs at high frequency in South Asia: India, Pakistan, Afghanistan, and the Himalayas. The group also occurs at >3% in some Iranian populations and is present at >30% in Roma from Croatia and Hungary.” ref
R-M458 (R1a1a1b1a1)
“R-M458 is a mainly Slavic SNP, characterized by its own mutation, and was first called cluster N. Underhill et al. (2009) found it to be present in modern European populations roughly between the Rhine catchment and the Ural Mountains and traced it to “a founder effect that … falls into the early Holocene period, 7.9±2.6 KYA.” M458 was found in one skeleton from a 14th-century grave field in Usedom, Mecklenburg-Vorpommern, Germany. The paper by Underhill et al. (2009) also reports a surprisingly high frequency of M458 in some Northern Caucasian populations (for example 27.5% among Karachays and 23.5% among Balkars, 7.8% among Karanogays and 3.4% among Abazas).” ref
Ceramic Mesolithic
“Pottery was found in Xianrendong cave, China, dating to 20,000–10,000 years ago. In North-Eastern Europe, Siberia, and certain southern European and North African sites, a “ceramic Mesolithic” can be distinguished between c. 9,000 to 5,850 years ago. Russian archaeologists prefer to describe such pottery-making cultures as Neolithic, even though farming is absent. This pottery-making Mesolithic culture can be found peripheral to the sedentary Neolithic cultures. It created a distinctive type of pottery, with point or knob base and flared rims, manufactured by methods not used by the Neolithic farmers. Though each area of Mesolithic ceramic developed an individual style, common features suggest a single point of origin. The earliest manifestation of this type of pottery may be in the region around Lake Baikal in Siberia. It appears in the Yelshanka culture on the Volga in Russia 9,000 years ago, and from there spread via the Dnieper-Donets culture to the Narva culture of the Eastern Baltic.” ref
“Spreading westward along the coastline it is found in the Ertebølle culture of Denmark and Ellerbek of Northern Germany, and the related Swifterbant culture of the Low Countries. A 2012 publication in the Science journal, announced that the earliest pottery yet known anywhere in the world was found in Xianrendong cave in China, dating by radiocarbon to between 20,000 and 19,000 years before present, at the end of the Last Glacial Period. The carbon 14 datation was established by carefully dating surrounding sediments. Many of the pottery fragments had scorch marks, suggesting that the pottery was used for cooking. These early pottery containers were made well before the invention of agriculture (dated to 10,000 to 8,000 BCE), by mobile foragers who hunted and gathered their food during the Late Glacial Maximum.” ref
Neman culture (7,100 to 5,000 years ago)
“The archaeological Neman culture (German: Memel-Kultur) existed from about 5100 to the 3rd millennium BCE, starting in the Mesolithic and continued into the middle Neolithic. It was located in the upper basin of the Neman River (present-day northern Poland, southern Lithuania, western Belarus and Kaliningrad Oblast). In the north, the Neman culture bordered the Kunda culture during the Mesolithic and the Narva culture during the Neolithic. The Neolithic began with the appearance of pottery in mid 5th millennium BCE. The Semi-Neolithic Neman culture was a successor of the Mesolithic Neman culture. Most of flint tools are very similar between both cultures. A new widespread development was knives with sharpened and flared point.” ref
“Pottery of the Neman culture had pointed bottoms and was made of clay mixed with organic matter or crushed quartzite. Some latter examples have flat bottoms. The vessels were a bit narrower and curvier than of the Narva culture. They were decorated with a thin layer of white clay and rows of small imprints around the rim. The rest of the vessel had diagonal stripes forming a pattern of a fishnet or more rows of small imprints. Some pottery found in settlements of the Neman culture was made by Narva culture. Such phenomenon is explained by trade of flint, which Narva culture in the north did not have. Towards the end of the Neman culture, the pottery became more varied and exhibits influence of the Rzucewo culture: imprints made by a cord or resembling a fir. Eventually, the culture was overtaken by the Globular Amphora culture and the Corded Ware culture. ” ref

Sintashta culture
“The Sintashta culture is a Middle Bronze Age archaeological culture of the Southern Urals, dated to the period c. 2200–1900 BCE. It is the first phase of the Sintashta–Petrovka complex, c. 2200–1750 BCE or around 4,200 to 3,750 years ago. The culture is named after the Sintashta archaeological site, in Chelyabinsk Oblast, Russia, and spreads through Orenburg Oblast, Bashkortostan, and Northern Kazakhstan. The Sintashta culture is thought to represent an eastward migration of peoples from the Corded Ware culture.” ref
“Sintashta settlements are estimated to have a population of between 200 and 700 individuals with economies that “heavily exploited domesticated cattle, sheep, and goats alongside horses with occasional hunting of wild fauna”. Anthony (2007) assumes that probably the people of the Sintashta culture spoke “Common-Indo-Iranian”. This identification is based primarily on similarities between sections of the Rig Veda, a religious text which includes ancient Indo-Iranian hymns recorded in Vedic Sanskrit, and the funerary rituals of the Sintashta culture as revealed by archaeology.” ref
“Some cultural similarities with Sintashta have also been found to be common with the Nordic Bronze Age of Scandinavia. There is linguistic evidence of interaction between Finno-Ugric and Indo-Iranian languages, showing influences from the Indo-Iranians into the Finno-Ugric culture. From the Sintashta culture the Indo-Iranian followed the migrations of the Indo-Iranians to Anatolia, the Iranian plateau and the Indian subcontintinent. From the 9th century BCE onward, Iranian languages also migrated westward with the Scythians back to the Pontic steppe where the proto-Indo-Europeans came from.” ref
“It is widely regarded as the origin of the Indo-Iranian languages (Indo-Iranic languages), whose speakers originally referred to themselves as the Arya. The earliest known chariots have been found in Sintashta burials, and the culture is considered a strong candidate for the origin of the technology, which spread throughout the Old World and played an important role in ancient warfare. Sintashta settlements are also remarkable for the intensity of copper mining and bronze metallurgy carried out there, which is unusual for a steppe culture. Among the main features of the Sintashta culture are high levels of militarism and extensive fortified settlements, of which 23 are known.” ref
“Because of the difficulty of identifying the remains of Sintashta sites beneath those of later settlements, the culture was only distinguished in the 1990s from the Andronovo culture. It was then recognised as a distinct entity, forming part of the “Andronovo horizon”. Koryakova (1998) concluded from their archaeological findings that the Sintashta culture originated from the interaction of the two precursors Poltavka culture and Abashevo culture. Allentoft et al. (2015) concluded from their genetic results that the Sintashta culture should have emerged from an eastward migration of peoples from the Corded Ware culture. In addition, Narasimshan et al. (2019) cautiously cite that “morphological data has been interpreted as suggesting that both Fedorovka and Alakul’ skeletons are similar to Sintashta groups, which in turn may reflect admixture of Neolithic forest HGs and steppe pastoralists, descendants of the Catacomb and Poltavka cultures.” ref
“Sintashta emerged during a period of climatic change that saw the already arid Kazakh steppe region become even colder and drier. The marshy lowlands around the Ural and upper Tobol rivers, previously favored as winter refuges, became increasingly important for survival. Under these pressures both Poltavka and Abashevo herders settled permanently in river valley strongholds, eschewing more defensible hill-top locations. Its immediate predecessor in the Ural-Tobol steppe was the Poltavka culture, an offshoot of the cattle-herding Yamnaya horizon that moved east into the region between 2800 and 2600 BCE. Several Sintashta towns were built over older Poltavka settlements or close to Poltavka cemeteries, and Poltavka motifs are common on Sintashta pottery.” ref
“Sintashta material culture also shows the influence of the late Abashevo culture, derived from the Fatyanovo-Balanovo culture, a collection of Corded Ware settlements in the forest steppe zone north of the Sintashta region that were also predominantly pastoralist. Radiocarbon dating indicates that the Sintashta culture dates to between c. 2200 and 1750 BCE, roughly contemporary with the associated Abashevo and Petrovka cultures. Some authors date the Petrovka culture slightly later, from c. 1900 BCE.” ref
“In Cis-Urals, burial sites Berezovaya and Tanabergen II showed Sintashta culture established there c. 2290–1750 BCE (68.2% probability), and the earliest values of this culture, in Trans-Urals, at the burial sites Sintashta II and Kamenny Ambar-5 (Kurgan 2) are c. 2200–2000 BCE. Chariots appear in southern Trans-Urals region in middle and late phases of the culture, c. 2050-1750 BC. Blöcher et al. (2023) consider Sintashta-Petrovka period came to an end in Trans-Urals c. 1900–1800 BCE.” ref
Genetics
“Allentoft et al. 2015 analyzed the remains of four individuals ascribed to the Sintastha culture. One male carried Y-haplogroup R1a and mt-J1c1b1a, while the other carried Y-R1a1a1b and mt-J2b1a2a. The two females carried U2e1e and U2e1h respectively. The study found a close autosomal genetic relationship between peoples of Corded Ware culture and Sintashta culture, which “suggests similar genetic sources of the two,” and may imply that “the Sintashta derives directly from an eastward migration of Corded Ware peoples.” ref
“Sintashta individuals and Corded Ware individuals both had a relatively higher ancestry proportion derived from the Central Europe, and both differed markedly in such ancestry from the population of the Yamnaya Culture and most individuals of the Poltavka Culture that preceded Sintashta in the same geographic region. Individuals from the Bell Beaker culture, the Unetice culture, and contemporary Scandinavian cultures were also found to be closely genetically related to Corded Ware. A particularly high lactose tolerance was found among Corded Ware and the closely related Nordic Bronze Age. In addition, the study found samples from the Sintashta culture to be closely genetically related to the succeeding Andronovo culture.” ref
“Narasimhan et al. 2019 analyzed the remains of several individuals associated with the Sintashta culture. mtDNA was extracted from two females buried at the Petrovka settlement. They were found to be carrying subclades of U2 and U5. The remains of fifty individuals from the fortified Sintastha settlement of Kamennyi Ambar was analyzed. This was the largest sample of ancient DNA ever sampled from a single site. The Y-DNA from thirty males was extracted. Eighteen carried R1a and various subclades of it (particularly subclades of R1a1a1), five carried subclades of R1b (particularly subclades of R1b1a1a), two carried Q1a and a subclade of it, one carried I2a1a1a, and four carried unspecified R1 clades. The majority of mtDNA samples belonged to various subclades of U, while W, J, T, H and K also occurred. A Sintashta male buried at Samara was found to be carrying R1b1a1a2 and J1c1b1a.” ref
“The authors of the study found the majority of Sintashta people (ca. 80%) to be closely genetically related to the people of the Corded Ware culture, the Srubnaya culture, the Potapovka culture, and the Andronovo culture. These were found to harbor mixed ancestry from the Yamnaya culture and peoples of the Central European Middle Neolithic, like the Globular Amphora culture. The remaining sampled Sintashta individuals belonged to various ancestral types different from the majority population, with affinities to earlier populations such as Eneolithic samples collected at Khvalynsk and hunter-gatherers from Tyumen Oblast in western Siberia. This indicates that the Sintashta settlement of Kamennyi Ambar was a cosmopolitan site that united a genetically heterogenous population in a single social group. Estimates based on DATES (Distribution of Ancestry Tracts of Evolutionary Signals) suggest that genetic characteristics typical of the Sintashta culture formed by c. 3200 BCE.” ref
Warfare
“The preceding Abashevo culture was already marked by endemic intertribal warfare; intensified by ecological stress and competition for resources in the Sintashta period. This drove the construction of fortifications on an unprecedented scale and innovations in military technique such as the invention of the war chariot. Increased competition between tribal groups may also explain the extravagant sacrifices seen in Sintashta burials, as rivals sought to outdo one another in acts of conspicuous consumption analogous to the North American potlatch tradition.” ref
“Sintashta artefact types such as spearheads, trilobed arrowheads, chisels, and large shaft-hole axes were taken east. Many Sintashta graves are furnished with weapons, although the composite bow associated later with chariotry does not appear. Higher-status grave goods include chariots, as well as axes, mace-heads, spearheads, and cheek-pieces. Sintashta sites have produced finds of horn and bone, interpreted as furniture (grips, arrow rests, bow ends, string loops) of bows; there is no indication that the bending parts of these bows included anything other than wood. Arrowheads are also found, made of stone or bone rather than metal. These arrows are short, 50–70 cm long, and the bows themselves may have been correspondingly short.” ref
“Sintashta culture, and the chariot, are also strongly associated with the ancestors of modern domestic horses, the DOM2 population. DOM2 horses originated from the Western Eurasia steppes, especially the lower Volga-Don, but not in Anatolia, during the late fourth and early third millennia BCE. Their genes may show selection for easier domestication and stronger backs.” ref
“The Sintashta economy came to revolve around copper metallurgy. Copper ores from nearby mines (such as Vorovskaya Yama) were taken to Sintashta settlements to be processed into copper and arsenical bronze. This occurred on an industrial scale: all the excavated buildings at the Sintashta sites of Sintashta, Arkaim and Ust’e contained the remains of smelting ovens and slag. Around 10% of graves, mostly adult male, contained artifacts related to bronze metallurgy (molds, ceramic nozzles, ore and slag remains, metal bars and drops). However, these metal-production related grave goods rarely co-occur with higher-status grave goods. This likely means that those who engaged in metal production were not at the top of the social-hierarchy, even though being buried at a cemetery evidences some sort of higher status.” ref
“Much of Sintashta metal was destined for export to the cities of the Bactria–Margiana Archaeological Complex (BMAC) in Central Asia. The metal trade between Sintashta and the BMAC for the first time connected the steppe region to the ancient urban civilisations of the Near East: the empires and city-states of modern Iran and Mesopotamia provided a large market for metals. These trade routes later became the vehicle through which horses, chariots and ultimately Indo-Iranian-speaking people entered the Near East from the steppe.” ref

Genetic history of Europe, Genetics and archaeogenetics of South Asia, and Genetic history of the Middle East
PIE Speakers and Haplogroups R1b as well as R1a
“According to three autosomal DNA studies, haplogroups R1b and R1a, now the most common in Europe (R1a is also very common in South Asia) would have expanded from the Pontic steppes, along with the Indo-European languages; they also detected an autosomal component present in modern Europeans which was not present in Neolithic Europeans, which would have been introduced with paternal lineages R1b and R1a, as well as Indo-European languages. Studies that analyzed ancient human remains in Ireland and Portugal suggest that R1b was introduced in these places along with autosomal DNA from the Pontic steppes.” ref
“The subclade R1a1a (R-M17 or R-M198) is most commonly associated with Indo-European speakers. Data so far collected indicate that there are two widely separated areas of high frequency, one in Eastern Europe, around Poland and the Russian core, and the other in South Asia, around Indo-Gangetic Plain. The historical and prehistoric possible reasons for this are the subject of on-going discussion and attention amongst population geneticists and genetic genealogists, and are considered to be of potential interest to linguists and archaeologists also. Ornella Semino et al. propose a postglacial (Holocene) spread of the R1a1 haplogroup from north of the Black Sea during the time of the Late Glacial Maximum, which was subsequently magnified by the expansion of the Kurgan culture into Europe and eastward.” ref
“A large, 2014 study by Underhill et al., using 16,244 individuals from over 126 populations from across Eurasia, concluded there was compelling evidence, that R1a-M420 originated in the vicinity of Iran. The mutations that characterize haplogroup R1a occurred ~10,000 years ago. Its defining mutation (M17) occurred about 10,000 to 14,000 years ago. Pamjav et al. (2012) believe that R1a originated and initially diversified either within the Eurasian Steppes or the Middle East and Caucasus region.” ref
Yamnaya culture
“All Yamnaya individuals sampled by Haak et al. (2015) belonged to the Y-haplogroup R1b. According to Jones et al. (2015) and Haak et al. (2015), autosomal tests indicate that the Yamnaya-people were the result of admixture between “Eastern Hunter-Gatherers” from eastern Europe (EHG) and “Caucasus hunter-gatherers” (CHG). Each of those two populations contributed about half the Yamnaya DNA. According to co-author Dr. Andrea Manica of the University of Cambridge:
The question of where the Yamnaya come from has been something of a mystery up to now […] we can now answer that, as we’ve found that their genetic make-up is a mix of Eastern European hunter-gatherers and a population from this pocket of Caucasus hunter-gatherers who weathered much of the last Ice Age in apparent isolation.” ref
“Based on these findings and by equating the people of the Yamnaya culture with the Proto-Indo-Europeans, David W. Anthony (2019) suggests that the Proto-Indo-European language formed mainly from a base of languages spoken by Eastern European hunter-gathers with influences from languages of northern Caucasus hunter-gatherers, in addition to a possible later influence from the language of the Maikop culture to the south (which is hypothesized to have belonged to the North Caucasian family) in the later neolithic or Bronze Age involving little genetic impact.” ref
Eastern European hunter-gatherers
“According to Haak et al. (2015), “Eastern European hunter-gatherers” who inhabited Russia were a distinctive population of hunter-gatherers with high affinity to a ~24,000-year-old Siberian from the Mal’ta-Buret’ culture, or other, closely related Ancient North Eurasian (ANE) people from Siberia and to the Western Hunter-Gatherers (WHG). Remains of the “Eastern European hunter-gatherers” have been found in Mesolithic or early Neolithic sites in Karelia and Samara Oblast, Russia, and put under analysis. Three such hunter-gathering individuals of the male sex have had their DNA results published. Each was found to belong to a different Y-DNA haplogroup: R1a, R1b, and J. R1b is also the most common Y-DNA haplogroup found among both the Yamnaya and modern-day Western Europeans. R1a is more common in Eastern Europeans and in the northern parts of the Indian subcontinent.” ref
Near East population
“The Near East population were most likely hunter-gatherers from the Caucasus (CHG) c.q. Iran Chalcolithic related people with a major CHG-component. Jones et al. (2015) analyzed genomes from males from western Georgia, in the Caucasus, from the Late Upper Palaeolithic (13,300 years old) and the Mesolithic (9,700 years old). These two males carried Y-DNA haplogroup: J* and J2a. The researchers found that these Caucasus hunters were probably the source of the farmer-like DNA in the Yamnaya, as the Caucasians were distantly related to the Middle Eastern people who introduced farming in Europe.” ref
“Their genomes showed that a continued mixture of the Caucasians with Middle Eastern took place up to 25,000 years ago, when the coldest period in the last Ice Age started. According to Lazaridis et al. (2016), “a population related to the people of the Iran Chalcolithic contributed ~43% of the ancestry of early Bronze Age populations of the steppe.” According to Lazaridis et al. (2016), these Iranian Chalcolithic people were a mixture of “the Neolithic people of western Iran, the Levant, and Caucasus Hunter-Gatherers.” Lazaridis et al. (2016) also note that farming spread at two places in the Near East, namely the Levant and Iran, from where it spread, Iranian people spreading to the steppe and south Asia.” ref
Northern and Central Europe
“Haak et al. (2015) studied DNA from 94 skeletons from Europe and Russia aged between 3,000 and 8,000 years old. They concluded that about 4,500 years ago there was a major influx into Europe of Yamnaya culture people originating from the Pontic–Caspian steppe north of the Black Sea and that the DNA of copper-age Europeans matched that of the Yamnaya. The four Corded Ware people could trace an astonishing three-quarters of their ancestry to the Yamnaya, according to the paper. That suggests a massive migration of Yamnaya people from their steppe homeland into Eastern Europe about 4500 years ago when the Corded Ware culture began, perhaps carrying an early form of Indo-European language.” ref
Bronze age Greece
“A 2017 archaeogenetics study of Mycenaean and Minoan remains published in the journal Nature concluded that the Mycenaean Greeks were genetically closely related with the Minoans but unlike the Minoans also had a 13-18% genetic contribution from Bronze Age steppe populations.” ref
Haplogroup R1a
“Haplogroup R1a, or haplogroup R-M420, is a human Y-chromosome DNA haplogroup that is distributed in a large region in Eurasia, extending from Scandinavia and Central Europe to southern Siberia and South Asia. While R1a originated ca. 22,000 to 25,000 years ago, its subclade M417 (R1a1a1) diversified ca. 5,800 years ago. The place of origin of the subclade plays a role in the debate about the origins of Proto-Indo-Europeans.” ref
“The split of R1a (M420) is computed to ca. 22,000 or 25,000 years ago, which is the time of the last glacial maximum. A 2014 study by Peter A. Underhill et al., using 16,244 individuals from over 126 populations from across Eurasia, concluded that there was “a compelling case for the Middle East, possibly near present-day Iran, as the geographic origin of hg R1a.” The ancient DNA record has shown the first R1a during the Mesolithic in Eastern Hunter-Gatherers (from Eastern Europe), and the earliest case of R* among Upper Paleolithic Ancient North Eurasians, from which the Eastern Hunter-Gatherers predominantly derive their ancestry. No early samples of R1a have so far been found in Iran.” ref
“According to Underhill et al. (2014), the downstream R1a-M417 subclade diversified into Z282 and Z93 circa 5,800 years ago “in the vicinity of Iran and Eastern Turkey.” Even though R1a occurs as a Y-chromosome haplogroup among various languages such as Slavic and Indo-Iranian, the question of the origins of R1a1a is relevant to the ongoing debate concerning the urheimat of the Proto-Indo-European people, and may also be relevant to the origins of the Indus Valley Civilization. R1a shows a strong correlation with Indo-European languages of Southern and Western Asia, Central and Eastern Europe and to some extent Scandinavia being most prevalent in Eastern Europe, West Asia, and South Asia. In Europe, Z282 is prevalent, particularly while in Asia Z93 dominates. The connection between Y-DNA R-M17 and the spread of Indo-European languages was first noted by T. Zerjal and colleagues in 1999.” ref
Proposed steppe dispersal of R1a1a: Indo-European migrations and Indo-Aryan migrations and R1a
“Semino et al. (2000) proposed Ukrainian origins, and a postglacial spread of the R1a1 gene during the Late Glacial Maximum, subsequently magnified by the expansion of the Kurgan culture into Europe and eastward. Spencer Wells proposes Central Asian origins, suggesting that the distribution and age of R1a1 points to an ancient migration corresponding to the spread by the Kurgan people in their expansion from the Eurasian steppe. According to Pamjav et al. (2012), R1a1a diversified in the Eurasian Steppes or the Middle East and Caucasus region:
Inner and Central Asia is an overlap zone for the R1a1-Z280 and R1a1-Z93 lineages [which] implies that an early differentiation zone of R1a1-M198 conceivably occurred somewhere within the Eurasian Steppes or the Middle East and Caucasus region as they lie between South Asia and Central- and Eastern Europe.” ref
“Three genetic studies in 2015 gave support to the Kurgan theory of Gimbutas regarding the Indo-European Urheimat. According to those studies, haplogroups R1b and R1a, now the most common in Europe (R1a is also common in South Asia) would have expanded from the Pontic–Caspian steppes, along with the Indo-European languages; they also detected an autosomal component present in modern Europeans which was not present in Neolithic Europeans, which would have been introduced with paternal lineages R1b and R1a, as well as Indo-European languages.” ref
Source of R1a1a1 in Corded Ware culture
“David Anthony considers the Yamnaya culture to be the Indo-European Urheimat. According to Haak et al. (2015), a massive migration from the Yamnaya culture northwards took place ca. 2,500 BCE or 4,622 years ago, accounting for 75% of the genetic ancestry of the Corded Ware culture, noting that R1a and R1b may have “spread into Europe from the East after 3,000 BCE” or 5,022 years ago. Yet, all their seven Yamnaya samples belonged to the R1b-M269 subclade, but no R1a1a has been found in their Yamnaya samples. This raises the question of where the R1a1a in the Corded Ware culture came from, if it was not from the Yamnaya culture.” ref
“Semenov & Bulat (2016) do argue for such an origin of R1a1a in the Corded Ware culture, noting that several publications point to the presence of R1a1 in the Comb Ware culture. Haak et al. (2015) found that part of the Yamnaya ancestry derived from the Middle East and that neolithic techniques probably arrived at the Yamnaya culture from the Balkans. The Rössen culture (4,600–4,300 BCE or 6,622-6,322 years ago), which was situated on Germany and predates the Corded Ware culture, an old subclade of R1a, namely L664, can still be found.” ref
Transcaucasia & West Asian origins and possible influence on Indus Valley Civilization
Kura–Araxes culture, Uruk period, and Origins of the Indus Valley Civilisation
“Part of the South Asian genetic ancestry derives from west Eurasian populations, and some researchers have implied that Z93 may have come to India via Iran and expanded there during the Indus Valley Civilization. However, according to Narasimhan et al. (2018), steppe pastoralists are a likely source for R1a in India.” ref
“Mascarenhas et al. (2015) proposed that the roots of Z93 lie in West Asia, and proposed that “Z93 and L342.2 expanded in a southeasterly direction from Transcaucasia into South Asia,” noting that such an expansion is compatible with “the archeological records of eastward expansion of West Asian populations in the 4th millennium BCE culminating in the so-called Kura-Araxes migrations in the post-Uruk IV period.” Yet, Lazaridis noted that sample I1635 of Lazaridis et al. (2016), their Armenian Kura-Araxes sample, carried Y-haplogroup R1b1-M415(xM269) (also called R1b1a1b-CTS3187).” ref
“According to Underhill et al. (2014) the diversification of Z93 and the “early urbanization within the Indus Valley […] occurred at [5,600 years ago] and the geographic distribution of R1a-M780 may reflect this.” Poznik et al. (2016) note that ‘striking expansions’ occurred within R1a-Z93 at ~4,500–4,000 years ago, which “predates by a few centuries the collapse of the Indus Valley Civilisation.” ref
Proposed South Asian origins
“Kivisild et al. (2003) have proposed either South or West Asia, while Mirabal et al. (2009) see support for both South and Central Asia. Sharma et al.(2009) showcased the existence of R1a in India beyond 18,000 years to possibly 44,000 years in origin. South Asian populations have the highest STR diversity within R1a1a, and subsequent older TMRCA datings, and R1a1a is present among both higher (Brahmin) castes and lower castes, although the frequency is higher among Brahmin castes. From these findings some researchers have concluded that R1a1a originated in South Asia, excluding a substantial genetic influx from Indo-European migrants.” ref
“However, this diversity, and the subsequent older TMRCA-datings, can also be explained by the historically high population numbers, which increases the likelihood of diversification and microsatellite variation. According to Sengupta et al. (2006), “[R1a1 and R2] could have actually arrived in southern India from a southwestern Asian source region multiple times.” Silva et al. (2017) noted that R1a in South Asia most “likely spread from a single Central Asian source pool, there do seem to be at least three and probably more R1a founder clades within the Subcontinent, consistent with multiple waves of arrival.” According to Martin P. Richards, co-author of Silva et al. (2017), “[the prevalence of R1a in India was] very powerful evidence for a substantial Bronze Age migration from central Asia that most likely brought Indo-European speakers to India.” ref
R-M458 (R1a1a1b1a1) 7,900 years old?
“R-M458 is a mainly Slavic SNP, characterized by its own mutation, and was first called cluster N. Underhill et al. (2009) found it to be present in modern European populations roughly between the Rhine catchment and the Ural Mountains and traced it to “a founder effect that […] falls into the early Holocene period, 7,900±2.6 KYA.” M458 was found in one skeleton from a 14th-century grave field in Usedom, Mecklenburg-Vorpommern, Germany. The paper by Underhill et al. (2009) also reports a surprisingly high frequency of M458 in some Northern Caucasian populations (for example 27.5% among Karachays and 23.5% among Balkars, 7.8% among Karanogays and 3.4% among Abazas).” ref
R-L260 (R1a1a1b1a1a)
“R1a1a1b1a1a (R-L260), commonly referred to as West Slavic or Polish, is a subclade of the larger parent group R-M458, and was first identified as an STR cluster by Pawlowski et al. 2002. In 2010 it was verified to be a haplogroup identified by its own mutation (SNP). It apparently accounts for about 8% of Polish men, making it the most common subclade in Poland. Outside of Poland it is less common. In addition to Poland, it is mainly found in the Czech Republic and Slovakia, and is considered “clearly West Slavic.” The founding ancestor of R-L260 is estimated to have lived between 2000 and 3000 years ago, i.e. during the Iron Age, with significant population expansion less than 1,500 years ago.” ref
“In Mesolithic Europe, R1a is characteristic of Eastern Hunter-Gatherers (EHGs). A male EHG of the Veretye culture buried at Peschanitsa near Lake Lacha in Arkhangelsk Oblast, Russia ca. 10,700 BCE or 12,722 years ago was found to be a carrier of the paternal haplogroup R1a5-YP1301 and the maternal haplogroup U4a. A Mesolithic male from Karelia ca. 8,800 to 7950 BCE or 10,822-9,972 years ago has been found to be carrying haplogroup R1a. A Mesolithic male buried at Deriivka ca. 7000 to 6700 BCE or 9,022-8,722 years ago carried the paternal haplogroup R1a and the maternal U5a2a. Another male from Karelia from ca. 5,500 to 5,000 BCE or 7,522-7,022 years ago, who was considered an EHG, carried haplogroup R1a. A male from the Comb Ceramic culture in Kudruküla ca. 5,900 to 3,800 BCE 7,922-5,822 years ago has been determined to be a carrier of R1a and the maternal U2e1.” ref
“According to archaeologist David Anthony, the paternal R1a-Z93 was found at Alexandria, Ukraine ca. 4000 BCE or 6,022 years ago, Sredny Stog culture, “the earliest known sample to show the genetic adaptation to lactase persistence (I3910-T).” R1a has been found in the Corded Ware culture, in which it is predominant. Examined males of the Bronze Age Fatyanovo culture belong entirely to R1a, specifically subclade R1a-Z93. Haplogroup R1a has later been found in ancient fossils associated with the Urnfield culture; as well as the burial of the remains of the Sintashta, Andronovo, the Pazyryk, Tagar, Tashtyk, and Srubnaya cultures, the inhabitants of ancient Tanais, in the Tarim mummies, and the aristocracy Xiongnu. The skeletal remains of a father and his two sons, from an archaeological site discovered in 2005 near Eulau (in Saxony-Anhalt, Germany) and dated to about 2600 BCE or 4,622 years ago, tested positive for the Y-SNP marker SRY10831.2. The Ysearch number for the Eulau remains is 2C46S. The ancestral clade was thus present in Europe at least 4600 years ago, in association with one site of the widespread Corded Ware culture.” ref
R1a and Europe
“In Europe, the R1a1 sub-clade is found at highest levels among peoples of Central and Eastern European descent, with results ranging from 35-65% among Czechs, Hungarians, Poles, Slovaks, western Ukrainians (particularly Rusyns), Belarusians, Moldovans, and Russians. In the Baltics, R1a1a frequencies decrease from Lithuania (45%) to Estonia (around 30%). There is a significant presence in peoples of Scandinavian descent, with the highest levels in Norway and Iceland, where between 20 and 30% of men are in R1a1a. Vikings and Normans may have also carried the R1a1a lineage further out; accounting for at least part of the small presence in the British Isles, the Canary Islands, and Sicily. In East Germany, where Haplogroup R1a1a reaches a peak frequency in Rostock at a percentage of 31.3%, it averages between 20 and 30%.” ref
“In Southern Europe, R1a1a is not common, but significant levels have been found in pockets, such as in the Pas Valley in Northern Spain, areas of Venice, and Calabria in Italy. The Balkans shows wide variation between areas with significant levels of R1a1a, for example, 36–39% in Slovenia, 27%-34% in Croatia, and over 30% in Greek Macedonia, but less than 10% in Albania, Kosovo, and parts of Greece south of Olympus gorge. R1a is virtually composed only of the Z284 subclade in Scandinavia. In Slovenia, the main subclade is Z282 (Z280 and M458), although the Z284 subclade was found in one sample of a Slovenian. There is a negligible representation of Z93 in each region other than Turkey.” ref
“West Slavs and Hungarians are characterized by a high frequency of the subclade M458 and a low Z92, a subclade of Z280. Hundreds of Slovenian samples and Czechs lack the Z92 subclade of Z280, while Poles, Slovaks, Croats and Hungarians only show a very low frequency of Z92. The Balts, East Slavs, Serbs, Macedonians, Bulgarians, and Romanians demonstrate a ratio Z280>M458 and a high, up to a prevailing share of Z92. Balts and East Slavs have the same subclades and similar frequencies in a more detailed phylogeny of the subclades. The Russian geneticist Oleg Balanovsky speculated that there is a predominance of the assimilated pre-Slavic substrate in the genetics of East and West Slavic populations, according to him the common genetic structure that contrasts East Slavs and Balts from other populations may suggest the explanation that the pre-Slavic substrate of the East Slavs consisted most significantly of Baltic-speakers, which at one point predated the Slavs in the cultures of the Eurasian steppe according to archaeological and toponymic references.” ref
R1a and Central Asia
“Zerjal et al. (2002) found R1a1a in 64% of a sample of the Tajiks of Tajikistan and 63% of a sample of the Kyrgyz of Kyrgyzstan. Haber et al. (2012) found R1a1a-M17(xM458) in 26.0% (53/204) of a set of samples from Afghanistan, including 60% (3/5) of a sample of Nuristanis, 51.0% (25/49) of a sample of Pashtuns, 30.4% (17/56) of a sample of Tajiks, 17.6% (3/17) of a sample of Uzbeks, 6.7% (4/60) of a sample of Hazaras, and in the only sampled Turkmen individual.” ref
“Di Cristofaro et al. (2013) found R1a1a-M198/M17 in 56.3% (49/87) of a pair of samples of Pashtuns from Afghanistan (including 20/34 or 58.8% of a sample of Pashtuns from Baghlan and 29/53 or 54.7% of a sample of Pashtuns from Kunduz), 29.1% (37/127) of a pool of samples of Uzbeks from Afghanistan (including 28/94 or 29.8% of a sample of Uzbeks from Jawzjan, 8/28 or 28.6% of a sample of Uzbeks from Sar-e Pol, and 1/5 or 20% of a sample of Uzbeks from Balkh), 27.5% (39/142) of a pool of samples of Tajiks from Afghanistan (including 22/54 or 40.7% of a sample of Tajiks from Balkh, 9/35 or 25.7% of a sample of Tajiks from Takhar, 4/16 or 25.0% of a sample of Tajiks from Samangan, and 4/37 or 10.8% of a sample of Tajiks from Badakhshan), 16.2% (12/74) of a sample of Turkmens from Jawzjan, and 9.1% (7/77) of a pair of samples of Hazara from Afghanistan (including 7/69 or 10.1% of a sample of Hazara from Bamiyan and 0/8 or 0% of a sample of Hazara from Balkh).” ref
“Malyarchuk et al. (2013) found R1a1-SRY10831.2 in 30.0% (12/40) of a sample of Tajiks from Tajikistan. Ashirbekov et al. (2017) found R1a-M198 in 6.03% (78/1294) of a set of samples of Kazakhs from Kazakhstan. R1a-M198 was observed with greater than average frequency in the study’s samples of the following Kazakh tribes: 13/41 = 31.7% of a sample of Suan, 8/29 = 27.6% of a sample of Oshaqty, 6/30 = 20.0% of a sample of Qozha, 4/29 = 13.8% of a sample of Qypshaq, 1/8 = 12.5% of a sample of Tore, 9/86 = 10.5% of a sample of Jetyru, 4/50 = 8.0% of a sample of Argyn, 1/13 = 7.7% of a sample of Shanyshqyly, 8/122 = 6.6% of a sample of Alimuly, 3/46 = 6.5% of a sample of Alban. R1a-M198 also was observed in 5/42 = 11.9% of a sample of Kazakhs of unreported tribal affiliation.” ref
R1a and South Asia
“In South Asia, R1a1a has often been observed in a number of demographic groups. In India, high frequencies of this haplogroup are observed in West Bengal Brahmins (72%) to the east, Gujarat Lohanas (60%) to the west, Khatris (67%) in the north, and Iyengar Brahmins (31%) in the south. It has also been found in several South Indian Dravidian-speaking Adivasis including the Chenchu (26%) and the Valmikis of Andhra Pradesh, Kota (22.58%), and the Kallar of Tamil Nadu suggesting that R1a1a is widespread in Tribal Southern Indians. Besides these, studies show high percentages in regionally diverse groups such as Manipuris (50%) to the extreme North-East and among Punjabis (47%) to the extreme North West. In Pakistan, it is found at 71% among the Mohanna tribe in Sindh province to the south and 46% among the Baltis of Gilgit-Baltistan to the north. Among the Sinhalese of Sri Lanka, 23% were found to be R1a1a (R-SRY1532) positive. Hindus of Chitwan District in the Terai region Nepal show it at 69%.” ref
R1a and East Asia
“The frequency of R1a1a is comparatively low among some Turkic-speaking groups like Yakuts, yet levels are higher (19 to 28%) in certain Turkic or Mongolic-speaking groups of Northwestern China, such as the Bonan, Dongxiang, Salar, and Uyghurs. A Chinese paper published in 2018 found R1a-Z94 in 38.5% (15 / 39) of a sample of Keriyalik Uyghurs from Darya Boyi / Darya Boye Village, Yutian County, Xinjiang (于田县达里雅布依乡), R1a-Z93 in 28.9% (22/76) of a sample of Dolan Uyghurs from Horiqol township, Awat County, Xinjiang (阿瓦提县乌鲁却勒镇), and R1a-Z93 in 6.3% (4/64) of a sample of Loplik Uyghurs from Karquga / Qarchugha Village, Yuli County, Xinjiang (尉犁县喀尔曲尕乡). R1a(xZ93) was observed only in one of 76 Dolan Uyghurs. Note that Darya Boyi Village is located in a remote oasis formed by the Keriya River in the Taklamakan Desert. A 2011 Y-dna study found Y-dna R1a1 in 10% of a sample of southern Hui people from Yunnan, 1.6% of a sample of Tibetan people from Xizang (Tibet Autonomous Region), 1.6% of a sample of Xibe people from Xinjiang, 3.2% of a sample of northern Hui from Ningxia, 9.4% of a sample of Hazak (Kazakhs) from Xinjiang, and rates of 24.0%, 22.2%, 35.2%, 29.2% in 4 different samples of Uyghurs from Xinjiang, 9.1% in a sample of Mongols from Inner Mongolia, 10% of a sample of Northern Han Chinese from Gansu and 8.9% of a sample of Northern Han from western Henan. A different subclade of R1 was also found in 1.5% of a sample of northern Hui from Ningxia.” ref
“In the same study, there were no cases of R1a detected at all in 6 samples of Han Chinese in Yunnan, 1 sample of Han in Guangxi, 5 samples of Han in Guizhou, 2 samples of Han in Guangdong, 2 samples of Han in Fujian, 2 samples of Han in Zhejiang, 1 sample of Han in Shanghai, 1 samples of Han in Jiangxi, 2 samples of Han in Hunan, 1 sample of Han in Hubei, 2 samples of Han in Sichuan, 1 sample of Han in Chongqing, 3 samples of Han in Shandong, 5 samples of Han in Gansu, 3 samples of Han in Jilin and 2 samples of Han in Heilongjiang. T-M70, R-M207 (a subclade of R1a), Q-M242, L-M20, J-P209, I-M170, H-M69, G-M201, C5-M356 and E-SRY4064 collectively make up only 6.79% of the total male population of East Asia (from samples in North Korea and China). The vast majority of East Asia is N-M231, C-M130 except for C5-M356, D-M174, and O-M175 which is 92.87% of the population and are all East Eurasian male haplogroups. R-M207 (a subclade of R1a) came into East Asia via the north from the Central South Asia region (CSA) during paleolithic times in the post-glacial period, especially R1a1a. R1a1a in East Asia is an extremely ancient subclade from the Central Asia-South Asia region and is older than the Western Eurasian (European_ and Central Asian-South Asian (CSA) R1a1*-M17, rivaling the R1a1*-M17 of IWest India in age from testing on variations in STR. The Europe and West Asian R1a1*-M17 split into 7 subbranches only after R1a1 came to North-East Asia, indicating R1a1 in East Asia is an extremely ancient one dating back 15,370 years ago juding from variation in STR (predating the more recent Aryan and Indo-European expansions).” ref
“In a 2014 paper, R1a1a has been detected in 1.8% (2/110) of Chinese samples. These two samples (R-M17, R-M198, R-M434, R-M458 for both) belonged to Han individuals from Fujian and Shanxi provinces. 40% of Salars, 45.2% of Tajiks of Xinjiang, 54.3% of Dongxiang, 60.6% of Tatars, and 68.9% of Kyrgyz in Xinjiang in northwestern China tested in one sample had R1a1-M17. Bao’an (Bonan) had the most haplogroup diversity of 0.8946±0.0305 while the other ethnic minorities in northwestern China had a high haplogroup diversity like Central Asians, of 0.7602±0.0546. In Eastern Siberia, R1a1a is found among certain indigenous ethnic groups including Kamchatkans and Chukotkans, and peaking in Itel’man at 22%.” ref
R1a and West Asia
“R1a1a has been found in various forms, in most parts of Western Asia, in widely varying concentrations, from almost no presence in areas such as Jordan, to much higher levels in parts of Kuwait and Iran. The Shimar (Shammar) Bedouin tribe in Kuwait show the highest frequency in the Middle East at 43%. Wells 2001, noted that in the western part of the country, Iranians show low R1a1a levels, while males of eastern parts of Iran carried up to 35% R1a1a. Nasidze et al. 2004 found R1a1a in approximately 20% of Iranian males from the cities of Tehran and Isfahan. Regueiro 2006 in a study of Iran, noted much higher frequencies in the south than the north.” ref
“A newer study has found 20.3% R-M17* among Kurdish samples which were taken in the Kurdistan Province in western Iran, 19% among Azerbaijanis in West Azerbaijan, 9.7% among Mazandaranis in North Iran in the province of Mazandaran, 9.4% among Gilaks in province of Gilan, 12.8% among Persian and 17.6% among Zoroastrians in Yazd, 18.2% among Persians in Isfahan, 20.3% among Persians in Khorasan, 16.7% Afro-Iranians, 18.4% Qeshmi “Gheshmi”, 21.4% among Persian Bandari people in Hormozgan and 25% among the Baloch people in Sistan and Baluchestan Province.” ref
“Di Cristofaro et al. (2013) found haplogroup R1a in 9.68% (18/186) of a set of samples from Iran, though with a large variance ranging from 0% (0/18) in a sample of Iranians from Tehran to 25% (5/20) in a sample of Iranians from Khorasan and 27% (3/11) in a sample of Iranians of unknown provenance. All Iranian R1a individuals carried the M198 and M17 mutations except one individual in a sample of Iranians from Gilan (n=27), who was reported to belong to R1a-SRY1532.2(xM198, M17).” ref
“Malyarchuk et al. (2013) found R1a1-SRY10831.2 in 20.8% (16/77) of a sample of Persians collected in the provinces of Khorasan and Kerman in eastern Iran, but they did not find any member of this haplogroup in a sample of 25 Kurds collected in the province of Kermanshah in western Iran. Further to the north of these Western Asian regions, on the other hand, R1a1a levels start to increase in the Caucasus, once again in an uneven way. Several populations studied have shown no sign of R1a1a, while the highest levels so far discovered in the region appears to belong to speakers of the Karachay-Balkar language among whom about one-quarter of men tested so far are in haplogroup R1a1a.” ref

ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref
Proto-Indo-Europeans: Western Steppe Herders
“The Proto-Indo-Europeans are a hypothetical prehistoric population of Eurasia who spoke Proto-Indo-European (PIE), the ancestor of the Indo-European languages according to linguistic reconstruction. Knowledge of them comes chiefly from that linguistic reconstruction, along with material evidence from archaeology and archaeogenetics. The Proto-Indo-Europeans likely lived during the late Neolithic, or roughly the 4th millennium BCE. Mainstream scholarship places them in the Pontic–Caspian steppe zone in Eastern Europe (present-day Ukraine and southern Russia).” ref
“Some archaeologists would extend the time depth of PIE to the middle Neolithic (5500 to 4500 BCE or 7,522-6,522 years ago) or even the early Neolithic (7500 to 5500 BCE or 9,522-7,522 years ago), and suggest alternative location hypotheses. By the early second millennium BCE, descendants of the Proto-Indo-Europeans had reached far and wide across Eurasia, including Anatolia (Hittites), the Aegean (the linguistic ancestors of Mycenaean Greece), the north of Europe (Corded Ware culture), the edges of Central Asia (Yamnaya culture), and southern Siberia (Afanasievo culture).” ref
“While ‘Proto-Indo-Europeans’ is used in scholarship to designate the group of speakers associated with the reconstructed proto-language and culture, the term ‘Indo-Europeans’ may refer to any historical people that speak an Indo-European language. In the words of philologist Martin L. West, “If there was an Indo-European language, it follows that there was a people who spoke it: not a people in the sense of a nation, for they may never have formed a political unity, and not a people in any racial sense, for they may have been as genetically mixed as any modern population defined by language.” ref
“Using linguistic reconstruction from old Indo-European languages such as Latin and Sanskrit, hypothetical features of the Proto-Indo-European language are deduced. Assuming that these linguistic features reflect the culture and environment of the Proto-Indo-Europeans, the following cultural and environmental traits are widely proposed:
- pastoralism, including domesticated cattle, horses, and dogs
- agriculture and cereal cultivation, including technology commonly ascribed to late-Neolithic farming communities, e.g., the plow
- transportation by or across water
- the solid wheel, used for wagons, but not yet chariots with spoked wheels
- worship of a sky god, *Dyḗus Ph2tḗr (lit. “sky father”; > Vedic Sanskrit Dyáuṣ Pitṛ́, Ancient Greek Ζεύς (πατήρ) / Zeus (dyeus)), vocative *dyeu ph2ter (> Latin Iūpiter, Illyrian Deipaturos)
- oral heroic poetry or song lyrics that used stock phrases such as imperishable fame (*ḱléwos ń̥dʰgʷʰitom) and the wheel of the sun (*sh₂uens kʷekʷlos).
- a patrilineal kinship-system based on relationships between men” ref
“A 2016 phylogenetic analysis of Indo-European folktales found that one tale, The Smith and the Devil, could be confidently reconstructed to the Proto-Indo-European period. This story, found in contemporary Indo-European folktales from Scandinavia to India, describes a blacksmith who offers his soul to a malevolent being (commonly a devil in modern versions of the tale) in exchange for the ability to weld any kind of materials together. The blacksmith then uses his new ability to stick the devil to an immovable object (often a tree), thus avoiding his end of the bargain. According to the authors, the reconstruction of this folktale to PIE implies that the Proto-Indo-Europeans had metallurgy, which in turn “suggests a plausible context for the cultural evolution of a tale about a cunning smith who attains a superhuman level of mastery over his craft.” ref
“Researchers have made many attempts to identify particular prehistoric cultures with the Proto-Indo-European-speaking peoples, but all such theories remain speculative. The scholars of the 19th century who first tackled the question of the Indo-Europeans’ original homeland (also called Urheimat, from German), had essentially only linguistic evidence. They attempted a rough localization by reconstructing the names of plants and animals (importantly the beech and the salmon) as well as the culture and technology (a Bronze Age culture centered on animal husbandry and having domesticated the horse).” ref
“The scholarly opinions became basically divided between a European hypothesis, positing migration from Europe to Asia, and an Asian hypothesis, holding that the migration took place in the opposite direction. In the early 20th century, the question became associated with the expansion of a supposed “Aryan race“, a now-discredited theory promoted during the expansion of European empires and the rise of “scientific racism“. The question remains contentious within some flavors of ethnic nationalism (see also Indigenous Aryans).” ref
“A series of major advances occurred in the 1970s due to the convergence of several factors. First, the radiocarbon dating method (invented in 1949) had become sufficiently inexpensive to be applied on a mass scale. Through dendrochronology (tree-ring dating), pre-historians could calibrate radiocarbon dates to a much higher degree of accuracy. And finally, before the 1970s, parts of Eastern Europe and Central Asia had been off-limits to Western scholars, while non-Western archaeologists did not have access to publications in Western peer-reviewed journals.” ref
“The pioneering work of Marija Gimbutas, assisted by Colin Renfrew, at least partly addressed this problem by organizing expeditions and arranging for more academic collaboration between Western and non-Western scholars. The Kurgan hypothesis, as of 2017 the most widely held theory, depends on linguistic and archaeological evidence, but is not universally accepted. It suggests PIE origin in the Pontic–Caspian steppe during the Chalcolithic. A minority of scholars prefer the Anatolian hypothesis, suggesting an origin in Anatolia during the Neolithic. Other theories (Armenian hypothesis, Out of India theory, Paleolithic Continuity Theory, Balkan hypothesis) have only marginal scholarly support.” ref
“In regard to terminology, in the 19th and early 20th centuries, the term Aryan was used to refer to the Proto-Indo-Europeans and their descendants. However, Aryan more properly applies to the Indo-Iranians, the Indo-European branch that settled parts of the Middle East and South Asia, as only Indic and Iranian languages explicitly affirm the term as a self-designation referring to the entirety of their people, whereas the same Proto-Indo-European root (*aryo-) is the basis for Greek and Germanic word forms which seem only to denote the ruling elite of Proto-Indo-European (PIE) society.” ref
“In fact, the most accessible evidence available confirms only the existence of a common, but vague, socio-cultural designation of “nobility” associated with PIE society, such that Greek socio-cultural lexicon and Germanic proper names derived from this root remain insufficient to determine whether the concept was limited to the designation of an exclusive, socio-political elite, or whether it could possibly have been applied in the most inclusive sense to an inherent and ancestral “noble” quality which allegedly characterized all ethnic members of PIE society. Only the latter could have served as a true and universal self-designation for the Proto-Indo-European people.” ref
“By the early twentieth century, this term had come to be widely used in a racist context referring to a hypothesized white, blonde, and blue-eyed “master race” (Herrenrasse), culminating with the pogroms of the Nazis in Europe. Subsequently, the term Aryan as a general term for Indo-Europeans has been largely abandoned by scholars (though the term Indo-Aryan is still used to refer to the branch that settled in Southern Asia).” ref
Proto-Indo-European Urheimat hypotheses and Indo-European migrations
“According to some archaeologists, PIE speakers cannot be assumed to have been a single, identifiable people or tribe, but were a group of loosely related populations ancestral to the later, still partially prehistoric, Bronze Age Indo-Europeans. This view is held especially by those archaeologists who posit an original homeland of vast extent and immense time depth. However, this view is not shared by linguists, as proto-languages, like all languages before modern transport and communication, occupied small geographical areas over a limited time span, and were spoken by a set of close-knit communities—a tribe in the broad sense. Researchers have put forward a great variety of proposed locations for the first speakers of Proto-Indo-European. Few of these hypotheses have survived scrutiny by academic specialists in Indo-European studies sufficiently well to be included in modern academic debate.” ref


ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref
“The shaman is, above all, a connecting figure, bridging several worlds for his people, traveling between this world, the underworld, and the heavens. He transforms himself into an animal and talks with ghosts, the dead, the deities, and the ancestors. He dies and revives. He brings back knowledge from the shadow realm, thus linking his people to the spirits and places which were once mythically accessible to all.–anthropologist Barbara Meyerhoff” ref

ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref
Elshanka culture
“The Elshanka culture (Russian: Елшанская культура) was a Subneolithic or very early Neolithic culture that flourished in the middle Volga region in the 7th millennium BCE. The sites are mostly individual graves scattered along the Samara and Sok rivers. They revealed Europe’s oldest pottery. The culture extended along the Volga from Ulyanovsk Oblast in the north through the Samara Bend towards Khvalynsk Hills and the Buzuluk District in the south. No signs of permanent dwellings have been found. Elshanka people appear to have been hunters and fishermen who had seasonal settlements at the confluences of rivers. Most grave goods come from such settlements.” ref
“Elshanka is believed to be the source from which the art of pottery spread south and westward towards the Balkans (with one particularly important site being the Surskoy Island in the Dnieper Rapids where pottery was made from 6200 BCE to 5800 BCE). Elshanka pots, dated from 6700 BCE onwards, usually have simple ornaments, though some have none. They were made “of a clay-rich mud collected from the bottoms of stagnant ponds, formed by the coiling method and were baken in open fires at 450-600 degrees Celsius”. A man buried at Lebyazhinka IV (a site usually assigned to the Elshanka culture) had the Haplogroup R1b. I. Vasiliev and A. Vybornov, citing the similarity of pottery, assert that Elshanka people were the descendants of the Zarzian culture who had been ousted from Central Asia by progressive desertification.” ref
“Other researchers see Elshanka ceramic industry as a local attempt at reproducing Zarzian pots. A rapid cooling around 6200 BCE and influences from the Lower Volga region led the Elshanka culture to be succeeded by the Middle Volga culture (with more complex ceramic ornaments) which lasted until the 5th millennium BCE. It was succeeded in the region by the better known Samara culture. Linguist Asko Parpola (2022) associates the Elshanka culture with the Pre-Proto-Indo-European language, stating that Elshanka expanded northwards into the forest zone as the Kama culture, reflecting a migration of Pre-PIE speakers into the Pre-Proto-Uralic-speaking area and thus possibly explaining the Indo-Uralic linguistic parallels.” ref
Dnieper–Donets culture
“The Dnieper–Donets culture complex (DDCC) (ca. 5th—4th millennium BCE) is a Mesolithic and later Neolithic archaeological culture found north of the Black Sea and dating to ca. 5000-4200 BCE. It has many parallels with the Samara culture, and was succeeded by the Sredny Stog culture.” ref
“Dnieper-Donets pottery was initially pointed based, but in later phases flat-based wares emerge. Their pottery is completely different from those made by the nearby Cucuteni–Trypillia culture. The importance of pottery appears to have increased throughout the existence of the Dnieper–Donets culture, which implies a more sedentary lifestyle. The early use of typical point base pottery interrelates with other Mesolithic cultures that are peripheral to the expanse of the Neolithic farmer cultures. The special shape of this pottery has been related to transport by logboat in wetland areas. Especially related are Swifterbant in the Netherlands, Ellerbek and Ertebølle in Northern Germany and Scandinavia, “Ceramic Mesolithic” pottery of Belgium and Northern France (including non-Linear pottery such as La Hoguette, Bliquy, Villeneuve-Saint-Germain), the Roucadour culture in Southwest France and the river and lake areas of Northern Poland and Russia.” ref
“In accordance with the original Kurgan hypothesis, J. Mallory (1997) suggested that the Dnieper-Donets people were Pre–Indo-European-speakers who were absorbed by Proto-Indo-Europeans expanding westwards from steppe-lands further east. According to David W. Anthony, the Indo-European languages were initially spoken by EHGs living in Eastern Europe, such as the Dnieper-Donets people. He (2007) also argues that the Dnieper-Donets people almost certainly spoke a different language from the people of the Cucuteni–Trypillia culture.” ref
“The areas of the upper Dniester in which the Dnieper–Donets culture was situated have mostly Baltic river names. That and the close relationship between the Dnieper–Donets culture and contemporary cultures of northeast Europe have caused the Dnieper–Donets culture to be identified with the later Balts. The precise role of the culture and its language to the derivation of the Pontic-Caspian cultures, such as Sredny Stog and Yamnaya culture, is open to debate, but the display of recurrent traits points to longstanding mutual contacts or to underlying genetic relations.” ref
“The physical remains recovered from graves of the Dnieper–Donets culture have been classified as “Proto-Europoid“. They are predominantly characterized as late Cro-Magnons with large and more massive features than the gracile Mediterranean peoples of the Balkan Neolithic. Males averaged 172 cm in height, which is much taller than contemporary Neolithic populations. Its rugged physical traits are thought to have genetically influenced later Indo-European peoples. Physical anthropologists have pointed out similarities in the physical type of the Dnieper-Donets people with the Mesolithic peoples of Northern Europe.” ref
“The peoples of the neighboring Sredny Stog culture, which eventually succeeded the Dnieper–Donets culture, were of a more gracile appearance. The first archaeogenetic analysis involving DDCC individuals was published by Nikitin et al. in 2012. The authors reported mtDNA haplogroups of two individuals from the Mykilske (Nikols’skoye in Russian) and Yasynuvatka (Yasinovatka) DDCC cemeteries. Haplogroups of west Eurasian (H, U3, U5a1a) and east Eurasian (C, C4a) descent have been identified. The authors linked the appearance of east Eurasian haplogroups with potential influence from northern Lake Baikal area.” ref
“Mathieson et al. (2018) analyzed 32 individuals from three Eneolithic cemeteries at Deriivka, Vilnyanka and Vovnigi, which Anthony (2019a) ascribed to the Dnieper–Donets culture. These individuals belonged exclusively to the paternal haplogroups R and I (mostly R1b and I2), and almost exclusively to the maternal haplogroup U (mostly U5, U4 and U2). This suggests that the Dnieper-Donets people were “distinct, locally derived population” of mostly of Eastern Hunter-Gatherer (EHG) descent, with Western Hunter-Gatherer (WHG) admixture. The WHG admixture appears to have increased in the transition from the Mesolithic to the Neolithic.” ref
“Unlike the Yamnaya culture, whose genetic cluster is known as Western Steppe Herder (WSH), in the Dnieper–Donets culture no Caucasian Hunter-Gatherer (CHG) or Early European Farmer (EEF) ancestry has been detected . At the same time, several Eneolithic individuals from the Deriivka I cemetery carried Anatolian Neolithic Farmer (ANF) – derived, as well as WSH ancestry. At the Vilnyanka cemetery, all the males belong to the paternal haplogroup I, which is common among WHGs. David W. Anthony suggests that this influx of WHG ancestry might be the result of EEFs pushing WHGs out of their territories to the east, where WHG males might have mated with EHG females.” ref
“Dnieper-Donets males and Yamnaya males carry the same paternal haplogroups (R1b and I2a), suggesting that the CHG and EEF admixture among the Yamnaya came through EHG and WHG males mixing with EEF and CHG females. According to Anthony, this suggests that the Indo-European languages were initially spoken by EHGs living in Eastern Europe. The Dnieper–Donets culture was succeeded by the Sredny Stog culture, its eastern neighbor, with whom it co-existed for a time before being finally absorbed. The Dnieper–Donets culture and the Sredny Stog culture were in turn succeeded by the Yamnaya culture. The Mikhaylovka culture, the Novodanilovka group and the Kemi Oba culture displays evidence of continuity from the Dnieper–Donets culture.” ref
“The Dnieper–Donets culture was originally a hunter-gatherer culture. The economic evidence from the earliest stages is almost exclusively from hunting and fishing. Among the sources of food hunted and foraged by the Dnieper-Donets people were aurochs, elk, red deer, roe, wild boar, fox, wildcat, hare, bear and onager. Their diet was primarily high protein, with meat, fish and nuts being consumed. From around 5200 BCE, the Dnieper-Donets people began keeping cattle, sheep and goats. Other domestic animals kept included pigs, horses and dogs.” ref
“During the following centuries, domestic animals from the Dnieper further and further east towards the Volga–Ural steppes, where they appeared ca. 4700-4600 BCE. Some scholars suggest that from about 4200 BCE, the Dnieper–Donets culture adopted agriculture. Domestic plants that have been recovered include millet, wheat and pea. At the same time, recent evidence suggests that millet did not arrive in west Eurasia until the Bronze Age. Evidence from skeletal remains suggest that plants were consumed. At the same time, systematic evidence of producing economy in DDCC is currently lacking. The presence of exotic goods in Dnieper-Donets graves indicates exchange relationships with the Caucasus.” ref
“David Anthony (2007: 155) dated the beginning of the Dnieper–Donets culture I roughly between 5800/5200 BCE. It quickly expanded in all directions, eventually absorbing all other local Neolithic groups. By 5200 BCE the Dnieper–Donets culture II followed, which ended between 4400/4200 BCE. The Dnieper–Donets culture complex was defined by the Soviet archaeologist Dmytro Telehin (Dmitriy Telegin) on proposition of another archaeologist Valentyn Danylenko in 1956. At that time Dmytro Telehin worked at the Institute of Archaeology of the Academy of Sciences of the Ukrainian SSR (1952 – 1990). In 1967 Telehin defended his doctorate dissertation “Dnieper–Donets Neolithic culture. The Dnieper–Donets culture produced no female figurines.” ref
“The Dnieper–Donets culture was distributed in the steppe and forest-steppe areas north of the Black Sea. Throughout its existence, rapid population growth and an expansion towards the steppe is noticeable. There are parallels with the contemporaneous Samara culture to the north. Striking similarities with the Khvalynsk culture and the Sredny Stog culture have also been detected. A much larger horizon from the upper Vistula to the lower half of Dnieper to the mid-to-lower Volga has therefore been drawn. Influences from the DDCC and the Sredny Stog culture on the Funnelbeaker culture have been suggested. An origin of the Funnelbeaker culture from the Dnieper–Donets culture has been suggested, but this is very controversial. The Dnieper–Donets culture was contemporary with the Bug–Dniester culture. It is clearly distinct from the Cucuteni–Trypillia culture.” ref
“The Dnieper–Donets culture is well known for about thirty of its cemeteries that have been discovered. This includes several large collective cemeteries of the Mariupol type. These contain around 800 individuals. It is evident that funerals were complex events that had several phases. Burials are mostly in large pits where the deceased were periodically placed and covered with ocher. In some cases, the deceased may have been exposed for a time before their bones were collected and buried. In most cases, however, the deceased were buried in the flesh without exposure. Deceased Dnieper-Donets people sometimes had only their skulls buried, but most often the entire bodies. The variants of Dnieper-Donets burial often appear in the same pits. Animal bones has also been found in the graves.” ref
“Certain Dnieper-Donets burials are accompanied with copper, crystal or porphyry ornaments, shell beads, bird-stone tubes, polished stone maces or ornamental plaques made of boar’s tusk. The items, along with the presence of animal bones and sophisticated burial methods, appear to have been a symbol of power. Certain deceased children were buried with such items, which indicates that wealth was inherited in Dnieper-Donets society. Very similar boar-tusk plaques and copper ornaments have been found at contemporary graves of the Samara culture in the middle Volga area. Maces of a different type than those of Dnieper-Donets have also been found. The wide adoption of such a status symbol attests to the existence of the institute of power in DDCC.” ref
“Individual, double and triple burials have also been found at DDCC cemeteries. These have been attributed to the earlier period of DDCC. Radiocarbon dates confirm the earlier chronology of individual DDCC burials compared to collective graves in large pits. Dnieper–Donets burials have been found near the settlement of Deriivka, which is associated with the Sredny Stog culture. The Dnieper–Donets culture continued using Mesolothic technology, but later phases see the appearance of polished stone axes, later flint and the disappearance of microliths. These tools were sometimes deposited in graves.” ref
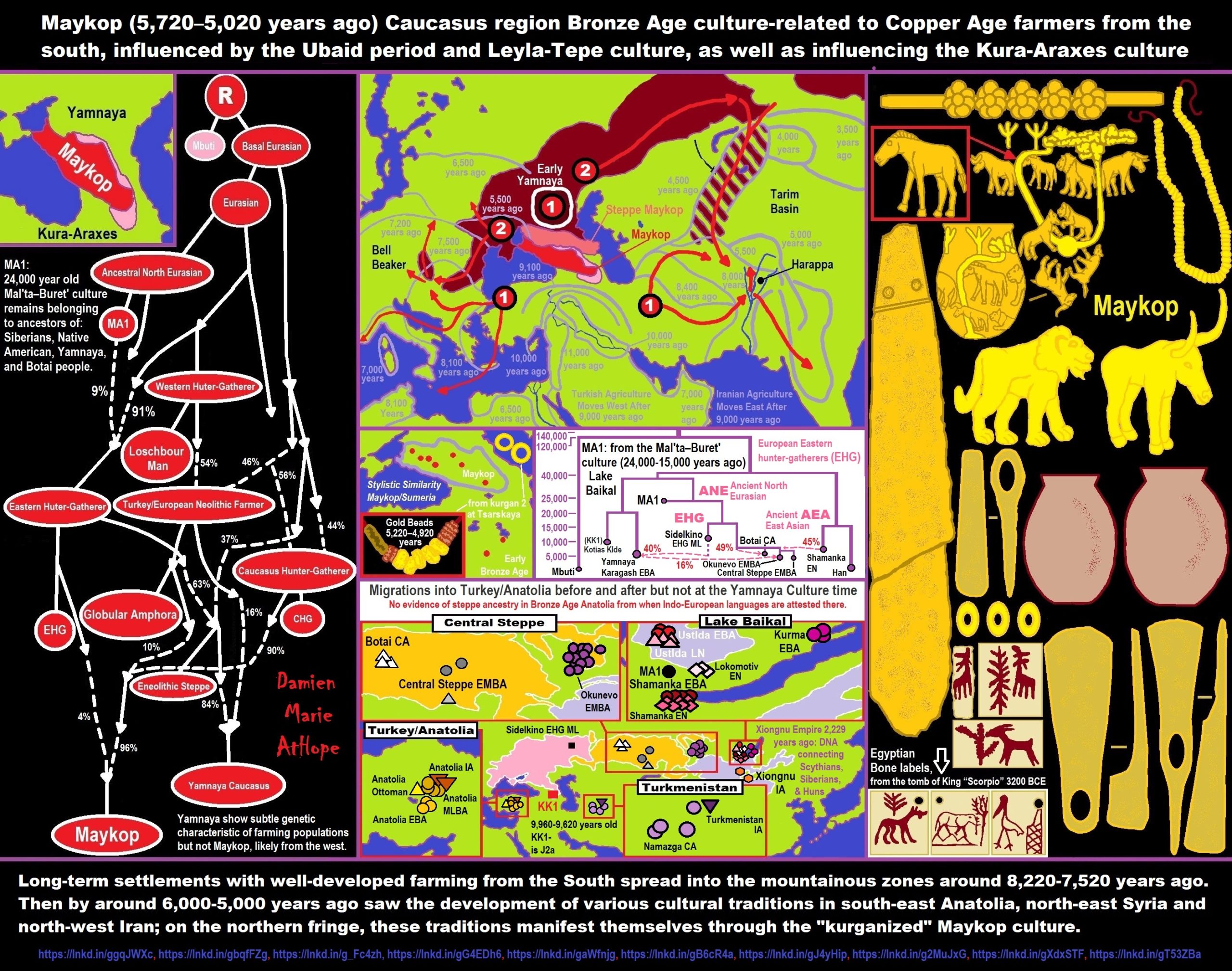
ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref
The Maykop culture, the R1b link to the Steppe?
“The Maykop culture (3700-2500 BCE) in the north-west Caucasus was culturally speaking a sort of southern extension of the Yamna horizon. Although not generally considered part of the Pontic-Caspian steppe culture due to its geography, the North Caucasus had close links with the steppes, as attested by numerous ceramics, gold, copper, and bronze weapons and jewelry in the contemporaneous cultures of Mikhaylovka, Sredny Stog, and Kemi Oba. The link between the northern Black Sea coast and the North Caucasus is older than the Maykop period. Its predecessor, the Svobodnoe culture (4400-3700 BCE), already had links to the Suvorovo-Novodanilovka and early Sredny Stog cultures. The even older Nalchik settlement (5000-4500 BCE) in the North Caucasus displayed a similar culture as Khvalynsk in the Caspian Steppe and Volga region. This may be the period when R1b started interacting and blending with the R1a population of the steppes.” ref
“The Yamna and Maykop people both used kurgan burials, placing their deads in a supine position with raised knees and oriented in a north-east/south-west axis. Graves were sprinkled with red ochre on the floor, and sacrificed domestic animal buried alongside humans. They also had in common horses, wagons, a heavily cattle-based economy with a minority of sheep kept for their wool, use of copper/bronze battle-axes (both hammer-axes and sleeved axes), and tanged daggers. In fact, the oldest wagons and bronze artefacts are found in the North Caucasus, and appear to have spread from there to the steppes.” ref
“Maykop was an advanced Bronze Age culture, actually one of the very first to develop metalworking, and therefore metal weapons. The world’s oldest sword was found at a late Maykop grave in Klady kurgan 31. Its style is reminiscent of the long Celtic swords, though less elaborated. Horse bones and depictions of horses already appear in early Maykop graves, suggesting that the Maykop culture might have been founded by steppe people or by people who had close link with them. However, the presence of cultural elements radically different from the steppe culture in some sites could mean that Maykop had a hybrid population. Without DNA testing it is impossible to say if these two populations were an Anatolian R1b group and a G2a Caucasian group, or whether R1a people had settled there too. The two or three ethnicities might even have cohabited side by side in different settlements. The one typical Caucasian Y-DNA lineage that does follow the pattern of Indo-European migrations is G2a-L13, which is found throughout Europe, Central Asia, and South Asia. In the Balkans, the Danube basin, and Central Europe its frequency is somewhat proportional to the percentage of R1b.” ref
“Maykop people are the ones credited for the introduction of primitive wheeled vehicles (wagons) from Mesopotamia to the Steppe. This would revolutionize the way of life in the steppe, and would later lead to the development of (horse-drawn) war chariots around 2000 BCE. Cavalry and chariots played an vital role in the subsequent Indo-European migrations, allowing them to move quickly and defeat easily anybody they encountered. Combined with advanced bronze weapons and their sea-based culture, the western branch (R1b) of the Indo-Europeans from the Black Sea shores are excellent candidates for being the mysterious Sea Peoples, who raided the eastern shores of the Mediterranean during the second millennium BCE.” ref
“The rise of the IE-speaking Hittites in Central Anatolia happened a few centuries after the disappearance of the Maykop and Yamna cultures. Considering that most Indo-European forms of R1b found in Anatolia today belong to the R1b-Z2103 subclade, it makes little doubt that the Hittites came to Anatolia via the Balkans, after Yamna/Maykop people invaded Southeast Europe. The Maykop and Yamna cultures were succeeded by the Srubna culture (1600-1200 BCE), possibly representing an advance of R1a-Z282 people from the northern steppes towards the Black Sea shores, filling the vacuum left by the R1b tribes who migrated to Southeast Europe and Anatolia.” ref
The Siberian & Central Asian branch corresponding to haplogroup R1b
“When R1b crossed the Caucasus in the Late Neolithic, it split into two main groups. The western one (L51) would settle the eastern and northern of the Black Sea. The eastern one (Z2103) migrated to the Don-Volga region, where horses were domesticated circa 4600 BCE. R1b probably mixed with indigenous R1a people and founded the Repin culture (3700-3300 BCE) a bit before the Yamna culture came into existence in the western Pontic Steppe. R1b would then have migrated with horses along the Great Eurasian Steppe until the Altai mountains in East-Central Asia, where they established the Afanasevo culture (c. 3600-2400 BCE). Afanasevo people might be the precursors of the Tocharian branch of Indo-European languages. In 2014, Clément Hollard of Strasbourg University tested three Y-DNA samples from the Afanasevo culture and all three turned out to belong to haplogroup R1b, including two to R1b-M269.” ref
“The R1b people who stayed in the Volga-Ural region were probably the initiators of the Poltavka culture (2700-2100 BCE), then became integrated into the R1a-dominant Sintashta-Petrovka culture (2100-1750 BCE) linked to the Indo-Aryan conquest of Central and South Asia (=> see R1a for more details). Nowadays in Russia R1b is found at higher frequencies among ethnic minorities of the Volga-Ural region (Udmurts, Komi, Mordvins, Tatars) than among Slavic Russians. R1b is also present in many Central Asian populations, the highest percentages being observed among the Uyghurs (20%) of Xinjiang in north-west China, the Yaghnobi people of Tajikistan (32%), and the Bashkirs (47%, or 62.5% in the Abzelilovsky district) of Bashkortostan in Russia (border of Kazakhstan). R1b-M73, found primarily in North Asia (Altai, Mongolia), Central Asia, and the North Caucasus is thought to have spread during the Neolithic from the Middle East to Central and North Asia, and therefore can be considered to be pre-Indo-European.” ref
The European & Middle Eastern branch corresponding to haplogroup R1b
“The Indo-Europeans’s bronze weapons and the extra mobility provided by horses would have given them a tremendous advantage over the autochthonous inhabitants of Europe, namely the native haplogroup C1a2, F, and I (descendants of Cro-Magnon) and the early Neolithic herders and farmers (G2a, H2, E1b1b, and T1a). This allowed R1a and R1b to replace most of the native male lineages (=> see How did R1b come to replace most of the older lineages in Western Europe?), although female lineages seem to have been less affected.” ref
“A comparison with the Indo-Iranian invasion of South Asia shows that 40% of the male linages of northern India are R1a, but less than 10% of the female lineages could be of Indo-European origin. The impact of the Indo-Europeans was more severe in Europe because European society 4,000 years ago was less developed in terms of agriculture, technology (no bronze weapons), and population density than that of the Indus Valley civilization. This is particularly true of the native Western European cultures where farming arrived much later than in the Balkans or Central Europe. Greece, the Balkans and the Carpathians were the most advanced of European societies at the time and were the least affected in terms of haplogroup replacement. neolithic lineages survived better in regions that were more difficult to reach or less hospitable to horse breeders, like the Alps, the Dinaric Alps, the Apennines, and Sardinia.” ref
The Conquest of “Old Europe” and Central Europe (4200-2500 BCE) corresponding to haplogroup R1b
“The first forays of Steppe people into the Balkans happened between 4200 BCE and 3900 BCE, when cattle herders equipped with horse-drawn wagons crossed the Dniester and Danube and apparently destroyed the towns of the Gumelnița, Varna, and Karanovo VI cultures in Eastern Romania and Bulgaria. A climatic change resulting in colder winters during this exact period probably pushed steppe herders to seek milder pastures for their stock, while failed crops would have led to famine and internal disturbance within the Danubian and Balkanic communities. The ensuing Cernavodă culture (Copper Age, 4000-3200 BCE), Coțofeni/Usatovo culture (Copper to Bronze Age, 3500-2500 BCE), Ezero culture (Bronze Age, 3300-2700 BCE), in modern Romania, seems to have had a mixed population of steppe immigrants and people from the old tell settlements. These Steppe immigrants were likely a mixture of both R1a and R1b lineages, with a probably higher percentage of R1a than later Yamna-era invasions.” ref
“The Steppe invaders would have forced many Danubian farmers to migrate to the Cucuteni-Trypillian towns in the eastern Carpathians, causing a population boom and a north-eastward expansion until the Dnieper valley, bringing Y-haplogroups G2a, I2a1 (probably the dominant lineage of the Cucuteni-Trypillian culture), E1b1b, J2a, and T1a in what is now central Ukraine. This precocious Indo-European advance westward was fairly limited, due to the absence of Bronze weapons and organized army at the time, and was indeed only possible thanks to climatic catastrophes which reduced the defenses of the towns of Old Europe. The Carphatian, Danubian, and Balkanic cultures were too densely populated and technologically advanced to allow for a massive migration.” ref
“In comparison, the forest-steppe R1a people successfully penetrated into the heart of Europe with little hindrance, due to the absence of developed agrarian societies around Poland and the Baltic. The Corded Ware culture (3200-1800 BCE) was a natural northern and western expansion of the Yamna culture, reaching as far west as Germany and as far north as Sweden and Norway. DNA analysis from the Corded Ware confirmed the presence of R1a and R1b in Poland c. 2700 BCE and R1a central Germany around 2600 BCE. The Corded Ware tribes expanded from the northern fringe of the Yamna culture where R1a lineages were prevalent over R1b ones.” ref
“The expansion of R1b people into Old Europe was slower, but proved inevitable. In 2800 BCE, by the time the Corded Ware had already reached Scandinavia, the Bronze Age R1b cultures had barely moved into the Pannonian Steppe. They established major settlements in the Great Hungarian Plain, the most similar habitat to their ancestral Pontic Steppes. Around 2500 BCE, the western branch of Indo-European R1b were poised for their next major expansion into modern Germany and Western Europe. By that time, the R1b immigrants had blended to a great extent with the indigenous Mesolithic and Neolithic populations of the Danubian basin, where they had now lived for 1,700 years.” ref
“The strongly patriarchal Indo-European elite remained almost exclusively R1b on the paternal side, but absorbed a high proportion of non-Indo-European maternal lineages. Hybridized, the new Proto-Indo-European R1b people would have lost most of their remaining Proto-Europoid or Mongolid features inherited from their Caspian origins (which were still clearly visible in numerous individuals from the Yamna period). Their light hair, eye and skin pigmentation, once interbred with the darker inhabitants of Old Europe, became more like that of modern Southern Europeans. The R1a people of the Corded Ware culture would come across far less populous societies in Northern Europe, mostly descended from the lighter Mesolithic population, and therefore retained more of their original pigmentation (although facial traits evolved considerably in Scandinavia, where the I1 inhabitants were strongly dolicocephalic and long-faced, as opposed to the brachycephalic and broad-faced Steppe people).” ref
The Conquest of Western Europe (2500-1200 BCE) corresponding to haplogroup R1b
“The R1b conquest of Europe happened in two phases. For nearly two millennia, starting from circa 4200 BCE, Steppe people limited their conquest to the rich Chalcolithic civilizations of the Carpathians and the Balkans. These societies possessed the world’s largest towns, notably the tell settlements of the Cucuteni-Tripolye culture. Nothing incited the R1b conquerors to move further into Western Europe at such an early stage, because most of the land north and west of the Alps was still sparsely populated woodland. The Neolithic did not reach the British Isles and Scandinavia before circa 4000 BCE. Even northern France and most of the Alpine region had been farming or herding for less than a millennium and were still quite primitive compared to Southeast Europe and the Middle East.” ref
“North-west Europe remained a tribal society of hunter-gatherers practicing only limited agriculture for centuries after the conquest of the Balkans by the Indo-Europeans. Why would our R1b “conquistadors” leave the comfort of the wealthy and populous Danubian civilizations for the harsh living conditions that lie beyond? Bronze Age people coveted tin, copper, and gold, of which the Balkans had plenty, but that no one had yet discovered in Western Europe.” ref
“R1b-L51 is thought to have arrived in Central Europe (Hungary, Austria, Bohemia) around 2500 BCE, approximately two millennia after the shift to the Neolithic lifestyle in these regions. Agrarian towns had started to develop. Gold and copper had begun to be mined. The prospects of a conquest were now far more appealing.” ref
“The archeological and genetic evidence (distribution of R1b subclades) point at several consecutive waves towards eastern and central Germany between 2800 BCE and 2300 BCE. The Unetice culture was probably the first culture in which R1b-L11 lineages played a major role. It is interesting to note that the Unetice period happen to correspond to the end of the Maykop (2500 BCE) and Kemi Oba (2200 BCE) cultures on the northern shores of the Black Sea, and their replacement by cultures descended from the northern steppes. It can therefore be envisaged that the (mostly) R1b population from the northern half of the Black Sea migrated westward due to pressure from other Indo-European people (R1a) from the north, for example that of the burgeoning Proto-Indo-Iranian branch, linked to the contemporary Poltavka and Abashevo cultures.” ref
“It is doubtful that the Bell Beaker culture (2900-1800 BCE) in Western Europe was already Indo-European because its attributes are in perfect continuity with the native Megalithic cultures. The Beaker phenomenon started during the Late Neolithic and Early Chalcolithic in Portugal and propagated to the north-east towards Germany. During the same period Bronze Age Steppe cultures spread from Germany in the opposite direction towards Iberia, France and Britain, progressively bringing R1b lineages into the Bell Beaker territory. It is more likely that the beakers and horses found across Western Europe during that period were the result of trade with neighboring Indo-European cultures, including the first wave of R1b into Central Europe. It is equally possible that the Beaker people were R1b merchants or explorers who traveled across Western Europe and brought back tales of riches poorly defended by Stone Age people waiting to be to be conquered. This would have prompted a full-scale Indo-European (R1b) invasion from about 2500 BCE in Germany, reaching the Atlantic (north of the Pyrenees at least) around 2200 BCE.” ref
“Ancient DNA tests conducted by Lee et al. (2012), Haak et al. (2015) and Allentoft et al. (2015) have all confirmed the presence of R1b-L51 (and deeper subclades such as P312 and U152) in Germany from the Bell Beaker period onwards, but none in earlier cultures. German Bell Beaker R1b samples only had about 50% of Yamna autosomal DNA and often possessed Neolithic non-Steppe mtDNA, which confirms that R1b invaders took local wives as they advanced westward. Another study by Olalde et al. (2017) confirmed that Iberian Bell Beakers were genetically distinct from the previously tested German samples. None of the Spanish or Portuguese individuals associated with Bell Beaker pottery possessed any Steppe admixture, and none belonged to the Indo-European haplogroup R1b-L23 or its subclades. Instead, they belonged to typical Megalithic lineages like G2a, I2a1, I2a2, and the Neolithic R1b-V88. The paper also confirmed a high frequency of R1b-L51 lineages in central Europe during the Beall Beaker period.” ref
In Britain, Megalithic individuals belonged exclusively to Y-haplogroup I2 (mostly I2a2 and I2a1b-L161), but were entirely replaced by R1b-L51 (mosly L21 clade) in the Early Bronze Age. This means that the Bell Beaker culture was not associated with one particular ethnic group. Beaker pottery originated in Megalithic Iberia, but then spread to France and central Europe and was used by invading R1b-L51 Steppe people, who brought it with them to the British Isles, while wiping out most of the indigenous Megalithic population. There was therefore no ‘Bell Beaker people’, but just various populations trading and using Beaker pots during that period.” ref
“DNA samples from the Unetice culture (2300-1600 BCE) in Germany, which emerged less than two centuries after the appearance of the first R1b-L51 individuals in the late Bell Beaker Germany, had a slightly higher percentage of Yamna ancestry (60~65%) and of Yamna-related mtDNA lineages, which indicates a migration of both Steppe men and women. That would explain why archeological artifacts from the Unetice culture are clearly Yamna-related (i.e. Indo-European), as they abruptly introduced new technologies and a radically different lifestyle, while the Bell Beaker culture was in direct continuity with previous Neolithic or Chalcolithic cultures. R1b men may simply have conquered the Bell Beaker people and overthrown the local rulers without obliterating the old culture due to their limited numbers. Taking the analogy of the Germanic migrations in the Late Antiquity, the R1b invasion of the Bell Beaker period was more alike to that of the Goths, Burgunds, and Vandals, who all migrated in small numbers, created new kingdoms within the Roman empire, but adopted Latin language and Roman culture. In contrast, the Corded Ware and Unetice culture involved large-scale migrations of Steppe people, who imposed their Indo-European language and culture and conquered people, just like the Anglo-Saxons or the Bavarians did in the 5th century.” ref
“The cultures that succeeded to Unetice in Central Europe, chronologically the Tumulus culture (1600-1200 BCE), Urnfield culture (1300-1200 BCE) and Hallstatt culture (1200-750 BCE) cultures remained typically Indo-European. The Hallstatt culture, centered around the Alps, is considered the first classical Celtic culture in Europe. It quickly expanded to France, Britain, Iberia, northern Italy, and the Danube valley, probably spreading for the first time Celtic languages, although not bronze technology nor R1b lineages, which had both already spread over much of western Europe during the Bell Beaker period. => See also Metal-mining and stockbreeding explain R1b dominance in Atlantic fringe” ref
Did the Indo-Europeans corresponding to haplogroup R1b really invade Western Europe?
“Proponents of the Paleolithic or Neolithic continuity model argue that bronze technology and horses could have been imported by Western Europeans from their Eastern European neighbors, and that no actual Indo-European invasion need be involved. It is harder to see how Italic, Celtic and Germanic languages were adopted by Western and Northern Europeans without at least a small scale invasion. It has been suggested that Indo-European (IE) languages simply disseminated through contact, just like technologies, or because it was the language of a small elite and therefore its adoption conferred a certain perceived prestige. However, people don’t just change language like that because it sounds nicer or more prestigious. Even nowadays, with textbooks, dictionaries, compulsory language courses at school, private language schools for adults, and multilingual TV programs, the majority of the people cannot become fluent in a completely foreign language, belonging to a different language family. The linguistic gap between pre-IE vernaculars and IE languages was about as big as between modern English and Chinese. English, Greek, Russian, and Hindi are all related IE languages and therefore easier to learn for IE speakers than non-IE languages like Chinese, Arabic, or Hungarian. From a linguistic point of view, only a wide-scale migration of IE speakers could explain the thorough adoption of IE languages in Western Europe – leaving only Basque as a remnant of the Neolithic languages.” ref
“One important archeological argument in favor of the replacement of Neolithic cultures by Indo-European culture in the Bronze Age comes from pottery styles. The sudden appearance of bronze technology in Western Europe coincides with ceramics suddenly becoming more simple and less decorated, just like in the Pontic Steppe. Until then, pottery had constantly evolved towards greater complexity and details for over 3,000 years. People do not just decide like that to revert to a more primitive style. Perhaps one isolated tribe might experiment with something simpler at one point, but what are the chances that distant cultures from Iberia, Gaul, Italy, and Britain all decide to undertake such an improbable shift around the same time? The best explanation is that this new style was imposed by foreign invaders. In this case, it is not mere speculation; there is ample evidence that this simpler pottery is characteristic of the steppes associated with the emergence of Proto-Indo-European speakers.” ref
“Besides pottery, archaeology provides ample evidence that the early Bronze Age in Central and Western Europe coincides with a radical shift in food production. Agriculture experiences an abrupt reduction in exchange for an increased emphasis on domesticates. This is also a period when horses become more common and cow milk is being consumed regularly. The overall change mimics the Steppic way of life almost perfectly. Even after the introduction of agriculture around 5200 BCE, the Bug-Dniester culture and later Steppe cultures were characterized by an economy dominated by herding, with only limited farming. This pattern expands into Europe exactly at the same time as bronze working.” ref
“Religious beliefs and arts undergo a complete reversal in Bronze Age Europe. Neolithic societies in the Near East and Europe had always worshipped female figurines as a form of fertility cult. The Steppe cultures, on the contrary, did not manufacture female figurines. As bronze technology spreads from the Danube valley to Western Europe, symbols of fertility and fecundity progressively disappear and are replaced by cultures of domesticated animals.” ref
“Another clue that Indo-European Steppe people came in great number to Central and Western Europe is to be found in burial practices. Neolithic Europeans either cremated their dead (e.g. Cucuteni-Tripolye culture) or buried them in collective graves (this was the case of Megalithic cultures). In the Steppe, each person was buried individually, and high-ranking graves were placed in a funeral chamber and topped by a circular mound. The body was typically accompanied by weapons (maces, axes, daggers), horse bones, and a dismantled wagon (or later chariot). These characteristic burial mounds are known as kurgans in the Pontic Steppe.” ref
“Men were given more sumptuous tombs than women, even among children, and differences in hierarchy are obvious between burials. The Indo-Europeans had a strongly hierarchical and patrilinear society, as opposed to the more egalitarian and matrilinear cultures of Old Europe. The proliferation of ststus-conscious male-dominant kurgans (or tumulus) in Central Europe during the Bronze Age is a clear sign that the ruling elite had now become Indo-European. The practice also spread to central Asia and southern Siberia, two regions where R1a and R1b lineages are found nowadays, just like in Central Europe.” ref
“The ceremony of burial is one of the most emotionally charged and personal aspect of a culture. It is highly doubtful that people would change their ancestral practice “just to do like the neighbors”. In fact, different funerary practices have co-existed side by side during the European Neolithic and Chalcolithic. The ascendancy of yet another constituent of the Pontic Steppe culture in the rest of Europe, and in this case one that does not change easily through contact with neighbors, adds up to the likelihood of a strong Indo-European migration. The adoption of some elements of a foreign culture tends to happen when one civilization overawes the adjacent cultures by its superiority.” ref
“This process is called ‘acculturation’. However, there is nothing that indicates that the Steppe culture was so culturally superior as to motivate a whole continent, even Atlantic cultures over 2000 km away from the Pontic Steppe, to abandon so many fundamental symbols of their own ancestral culture, and even their own language. In fact, Old Europe was far more refined in its pottery and jewelry than the rough Steppe people. The Indo-European superiority was cultural but military, thanks to horses, bronze weapons, and an ethic code valuing individual heroic feats in war (these ethic values are known from the old IE texts, like the Rig Veda, Avesta, or the Mycenaean and Hittite literature).” ref
“After linguistics and archaeology, the third category of evidence comes from genetics itself. It had first been hypothesized that R1b was native to Western Europe, because this is where it was most prevalent. It has since been proven that R1b haplotypes displayed higher microsatellite diversity in Anatolia and in the Caucasus than in Europe. European subclades are also more recent than Middle Eastern or Central Asian ones. The main European subclade, R-P312/S116, only dates back to approximately 3500 to 3000 BCE. It does not mean that the oldest common ancestor of this lineage arrived in Western Europe during this period, but that the first person who carried the mutation R-P312/S116 lived at least 5,000 years ago, assumably somewhere in the lower Danube valley or around the Black Sea. In any case, this timeframe is far too recent for a Paleolithic origin or a Neolithic arrival of R1b. The discovery of what was thought to be “European lineages” in Central Asia, Pakistan, and India hit the final nail on the coffin of a Paleolithic origin of R1b in Western Europe, and confirmed the Indo-European link.” ref
“All the elements concur in favor of a large scale migration of Indo-European speakers (possibly riding on horses) to Western Europe between 2500 to 2100 BCE, contributing to the replacement of the Neolithic or Chalcolithic lifestyle by an inherently new Bronze Age culture, with simpler pottery, less farming, more herding, new rituals (single graves) and new values (patrilinear society, warrior heroes) that did not evolve from local predecessors.” ref
The Atlantic Celtic branch (L21) corresponding to haplogroup R1b
“The Proto-Italo-Celto-Germanic R1b people had reached in what is now Germany by 2500 BCE. By 2300 BCE they had arrived in large numbers and founded the Unetice culture. Judging from the propagation of bronze working to Western Europe, those first Indo-Europeans reached France and the Low Countries by 2200 BCE, Britain by 2100 BCE and Ireland by 2000 BCE, and Iberia by 1800 BCE. This first wave of R1b presumably carried R1b-L21 lineages in great number (perhaps because of a founder effect), as these are found everywhere in western, northern, and Central Europe. Cassidy et al. (2015) confirmed the presence of R1b-L21 (DF13 and DF21 subclades) in Ireland around 2000 BCE. Those genomes closely resembled those of the Unetice culture autosomally, but differed greatly from the earlier Neolithic Irish samples. This confirms that a direct migration of R1b-L21 from Central Europe was responsible for the introduction of the Bronze Age to Ireland.” ref
“The early split of L21 from the main Proto-Celtic branch around Germany would explain why the Q-Celtic languages (Goidelic and Hispano-Celtic) diverged so much from the P-Celtic branch (La Tène, Gaulish, Brythonic), which appears to have expanded from the later Urnfield and Hallstat cultures. Some L21 lineages from the Netherlands and northern Germany later entered Scandinavia (from 1700 BCE) with the dominant subclade of the region, R1b-S21/U106 (see below). The stronger presence of L21 in Norway and Iceland can be attributed to the Norwegian Vikings, who had colonized parts of Scotland and Ireland and taken slaves among the native Celtic populations, whom they brought to their new colony of Iceland and back to Norway. Nowadays about 20% of all Icelandic male lineages are R1b-L21 of Scottish or Irish origin.” ref
“In France, R1b-L21 is mainly present in historical Brittany (including Mayenne and Vendée) and in Lower Normandy. This region was repopulated by massive immigration of insular Britons in the 5th century due to pressure from the invading Anglo-Saxons. However, it is possible that L21 was present in Armorica since the Bronze age or the Iron age given that the tribes of the Armorican Confederation of ancient Gaul already had a distinct identity from the other Gauls and had maintained close ties with the British Isles at least since the Atlantic Bronze Age.” ref
The Gallic & Iberian branch (DF27/S250) corresponding to haplogroup R1b
“The first Proto-Celtic R1b lineages to reach France and the Iberian peninsula from Central Europe were probably L21 and DF27. Whereas L21 might have taken a northern route through Belgium and northern France on its way to the British Isles, DF27 seems to have spread all over France but heading in greater number toward the south.” ref
“The Bronze Age did not appear in Iberia until 1800 BCE, and was mostly confined to the cultures of El Argar and Los Millares in south-east Spain, with sporadic sites showing up in Castile by 1700 BCE and in Extremadura and southern Portugal by 1500 BCE. These Early Bronze Age sites typically did not have more than some bronze daggers or axes and cannot be considered proper Bronze Age societies, but rather Copper Age societies with occasional bronze artifacts (perhaps imported). These cultures might have been founded by small groups of R1b adventurers looking for easy conquests in parts of Europe that did not yet have bronze weapons. They would have become a small ruling elite, would have had children with local women, and within a few generations their Indo-European language would have been lost, absorbed by the indigenous languages (=> see How did the Basques become R1b?).” ref
“Martiniano et al. (2017) sequenced the genomes of various skeletons from West Iberia dating from the Middle and Late Neolithic, Chalcolithic and Middle Bronze Age (since the Early Bronze Age did not reach that region). They found that Neolithic and Chalcolithic individuals belonged to Y-haplogroups I*, I2a1, and G2a. In contrast, all three Bronze Age Portuguese men tested belonged to R1b (one M269 and two P312), although they carried Neolithic Iberian maternal lineages (H1, U5b3, X2b) and lacked any discernible Steppe admixture. This is concordant with a scenario of Indo-European R1b men entering Iberia from 1800 BCE as a small group of adventurers and taking local wives, thus diluting their DNA at each generation, until hardly any Steppe admixture was left after a few centuries, by the time they reached Portugal. Nowadays, Spaniards and Portuguese do possess about 25% of Steppe admixture, which means that other more important Indo-European migrations took place later on, during the Late Bronze Age and the Iron Age.” ref
“Iberia did not become a fully-fledged Bronze Age society until the 13th century BCE, when the Urnfield culture (1300-1200 BCE) expanded from Germany to Catalonia via southern France, then the ensuing Hallstatt culture (1200-750 BCE) spread throughout most of the peninsula (especially the western half). This period belongs to the wider Atlantic Bronze Age (1300-700 BCE), when Iberia was connected to the rest of Western Europe through a complex trade network.” ref
“It is hard to say when exactly DF27 entered Iberia. Considering its overwhelming presence in the peninsula and in south-west France, it is likely that DF27 arrived early, during the 1800 to 1300 BCE period, and perhaps even earlier, if R1b adventurers penetrated the Bell Beaker culture, as they appear to have done all over Western Europe from 2300 BCE to 1800 BCE. The Atlantic Bronze Age could correspond to the period when DF27 radiated more evenly around Iberia and ended up, following Atlantic trade routes, all the way to the British Isles, the Netherlands, and Scandinavia.” ref
The Italo-Celtic branch (S28/U152/PF6570) corresponding to haplogroup R1b
“Furtwängler et al. (2020) analyzed 96 ancient genomes from Switzerland, Southern Germany, and the Alsace region in France, covering the Middle/Late Neolithic to Early Bronze Age. They confirmed that R1b arrived in the region during the transitory Bell Beaker period (2800-1800 BCE). The vast majority of Bell Beaker R1b samples belonged to the U152 > L2 clade (11 out of 14; the other being P312 or L51).” ref
“Starting circa 1300 BCE, a new Bronze Age culture flourished around the Alps thanks to the abundance of metal in the region, and laid the foundation for the classical Celtic culture. It was actually the succession of three closely linked culture: the Urnfield culture, which would evolve into the Hallstatt culture (from 1200 BCE) and eventually into the La Tène culture (from 450 BCE). After the Unetice expansion to Western Europe between 2300 and 1800 BCE, the Urnfield/Hallstatt/La Tène period represents the second major R1b expansion that took place from Central Europe, pushing west to the Atlantic, north to Scandinavia, east to the Danubian valley, and eventually as far away as Greece, Anatolia, Ukraine and Russia, perhaps even until the Tarim basin in north-west China (=> see Tarim mummies.” ref
“R1b-U152 would have entered Italy in successive waves from the northern side of the Alps, starting in 1700 BCE with the establishment of the Terramare culture in the Po Valley. From 1200 BCE, a larger group of Hallstatt-derived tribes founded the Villanova culture (see below). This is probably the migration that brought the Italic-speaking tribes to Italy, who would have belonged mainly the Z56 clade of R1b-U152. During the Iron Age, the expansion of the La Tène culture from Switzerland is associated with the diffusion of the Z36 branch, which would generate the Belgae around modern Belgium and in the Rhineland, the Gauls in France, and the Cisalpine Celts in Italy.” ref
“Antonio et al. (2019) analysed the genomes of Iron Age Latins dating between 900 and 200 BCE, and the samples tested belonged primarily to haplogroup R1b-U152 (including the clades L2, Z56 and Z193), as well as one R1b-Z2103 and one R1b-Z2118. One common linguistic trait between Italic and Gaulish/Brythonic Celtic languages linked to the Hallstatt expansion is that they shifted the original IE *kw sound into *p.” ref
They are known to linguists as the P-Celtic branch (as opposed to Q-Celtic). It is thought that this change occurred due to the inability to pronounce the *kw sound by the pre-Indo-European population of Central Europe, Gaul, and Italy, who were speakers of Afro-Asiatic dialects that had evolved from Near-Eastern languages inherited from the Neolithic. The Etruscans, although later incomers from the eastern Mediterranean, also fit in this category. It has recently been acknowledged that Celtic languages borrowed part of their grammar from Afro-Asiatic languages.” ref
“This shift could have happened when the Proto-Italo-Celtic speakers moved from the steppes to the Danube basin and mixed with the population of Near-Eastern farmers belonging to haplogroups E1b1b, G2a, J, and T. However, such an early shift would not explain why Q-Celtic and Germanic languages did not undergo the same linguistic mutation. It is therefore more plausible that the shift happened after the Proto-Italo-Celts and Proto-Germanics had first expanded across all western and northern Europe. The S28/U152 connection to P-Celtic (and Italic) suggests that the shift took place around the Alps after 1800 BCE, but before the invasion of Italy by the Italic tribes circa 1200 BCE.” ref
“The expansion of the Urnfield/Hallstatt culture to Italy is evident in the form of the Villanovan culture (c. 1100-700 BCE), which shared striking resemblances with the Urnfield/Hallstatt sites of Bavaria and Upper Austria. The Villanova culture marks a clean break with the previous Terramare culture. Although both cultures practiced cremation, whereas Terramare people placed cremated remains in communal ossuaries like their Neolithic ancestors from the Near East, Villanovans used distinctive Urnfield-style double-cone shaped funerary urns, and elite graves containing jewelry, bronze armor and horse harness fittings were separated from ordinary graves, showing for the first time the development of a highly hierarchical society, so characteristic of Indo-European cultures.” ref
“Quintessential Indo-European decorations, such as swastikas, also make their appearance. Originally a Bronze-age culture, the Villanova culture introduced iron working to the Italian peninsula around the same time as it appeared in the Hallstatt culture, further reinforcing the link between the two cultures. In all likelihood, the propagation of the Villanova culture represents the Italic colonization of the Italian peninsula. The highest proportion of R1b-U152 is found precisely where the Villanovans were the more strongly established, around modern Tuscany and Emilia-Romagna. The Villanova culture was succeeded by the Etruscan civilisation, which displayed both signs of continuity with Villanova and new hybrid elements of West Asian origins, probably brought by Anatolian settlers (who would have belonged to a blend of haplogroups G2a, J2, and R1b-Z2103).” ref
The Germanic branch (S21/U106/M405) corresponding to haplogroup R1b
“The principal Proto-Germanic branch of the Indo-European family tree is R1b-S21 (a.k.a. U106 or M405). This haplogroup is found at high concentrations in the Netherlands and north-west Germany. It is likely that R1b-S21 lineages expanded in this region through a founder effect during the Unetice period, then penetrated into Scandinavia around 1700 BCE (probably alongside R1a-L664), thus creating a new culture, that of the Nordic Bronze Age (1700-500 BCE). R1b-S21 would then have blended for more than a millennium with preexisting Scandinavian populations, represented by haplogroups I1, I2-L801, R1a-Z284. When the Germanic Iron Age started c. 500 BCE, the Scandinavian population had developed a truly Germanic culture and language, but was divided in many tribes with varying levels of each haplogroup. R1b-S21 became the dominant haplogroup among the West Germanic tribes, but remained in the minority against I1 and R1a in East Germanic and Nordic tribes, including those originating from Sweden such as the Goths, the Vandals, and Lombards.” ref
“The presence of R1b-S21 in other parts of Europe can be attributed almost exclusively to the Germanic migrations that took place between the 3rd and the 10th century. The Frisians and Anglo-Saxons disseminated this haplogroup to England and the Scottish Lowlands, the Franks to Belgium and France, the Burgundians to eastern France, the Suebi to Galicia, and northern Portugal, and the Lombards to Austria and Italy. The Goths help propagate S21 around Eastern Europe, but apparently their Germanic lineages were progressively diluted by blending with Slavic and Balkanic populations, and their impact in Italy, France, and Spain was very minor. Later the Danish and Norwegian Vikings have also contributed to the diffusion of R1b-S21 (alongside I1, I2b1, and R1a) around much of Western Europe, but mainly in Iceland, in the British Isles, in Normandy, and in the southern Italy.” ref
“From the Late Middle Ages until the early 20th century, the Germans expanded across much of modern Poland, pushing as far as Latvia to the north-east and Romania to the south-east. During the same period the Austrians built an empire comprising what is now the Czech Republic, Slovakia, Hungary, Slovenia, Croatia, Serbia, and parts of Romania, western Ukraine, and southern Poland. Many centuries of German and Austrian influence in central and Eastern Europe resulted in a small percentage of Germanic lineages being found among modern populations. In Romania 4% of the population still consider themselves German. The low percentage of R1b-S21 in Finland, Estonia, and Latvia can be attributed to the Swedish or Danish rule from the late Middle Ages to the late 19th century.” ref
“O’Sullivan et al. (2018) tested the genomes of Merovingian nobles from an early Medieval Alemannic graveyard in Baden-Württemberg. Apart from one individual belonging to haplogroup G2a2b1, all men were members of R1b, and all samples that yielded deep clade results fell under the R1b-U106 > Z381 > Z301 > L48 > Z9 > Z325 clade. The lineage of the Kings of France was inferred from the Y-DNA of several descendant branches (see famous members below) and also belongs to R1b-U106 > Z381. Their earliest-known male-line ancestor was from Robert II, Count of Hesbaye, a Frankish nobleman from present-day Belgium. The House of Wettin (see famous members below), one of the oldest dynasties in Europe, which ruled over many states at various times in history, was yet another well-known noble Germanic lineage part of R1b-U106 > Z381.” ref
How did R1b come to replace most of the older lineages in Western Europe?
“Until recently it was believed that R1b originated in Western Europe due to its strong presence in the region today. The theory was that R1b represented the Paleolithic Europeans (Cro-Magnon) that had sought refuge in the Franco-Cantabrian region at the peak of the last Ice Age, then recolonised Central and Northern Europe once the ice sheet receded. The phylogeny of R1b proved that this scenario was not possible, because older R1b clades were consistently found in Central Asia and the Middle East, and the youngest in Western and Northern Europe. There was a clear gradient from East to West tracing the migration of R1b people (see map above). This age of the main migration from the shores of the Black Sea to Central Europe also happened to match the timeframe of the Indo-European invasion of Europe, which coincides with the introduction of the Bronze-Age culture in Western Europe, and the proliferation of Italo-Celtic and Germanic languages.” ref
“Historians and archeologists have long argued whether the Indo-European migration was a massive invasion, or rather a cultural diffusion of language and technology spread only by a small number of incomers. The answer could well be “neither”. Proponents of the diffusion theory would have us think that R1b is native to Western Europe, and R1a alone represents the Indo-Europeans. The problem is that haplogroup R did arise in Central Asia, and R2 is still restricted to Central and South Asia, while R1a and the older subclades of R1b are also found in Central Asia. The age of R1b subclades in Europe coincides with the Bronze-Age. R1b must consequently have replaced most of the native Y-DNA lineages in Europe from the Bronze-Age onwards.” ref
“However, a massive migration and nearly complete annihilation of the Paleolithic population can hardly be envisaged. Western Europeans do look quite different in Ireland, Holland, Aquitaine, or Portugal, despite being all regions where R1b is dominant. Autosomal DNA studies have confirmed that the Western European population is far from homogeneous. A lot of maternal lineages (mtDNA) also appear to be of Paleolithic origin (e.g. H1, H3, U5, or V) based on ancient DNA tests. What a lot of people forget is that there is also no need of a large-scale exodus for patrilineal lineages to be replaced fairly quickly. Here is why.” ref
- “Polygamy. Unlike women, men are not limited in the number of children they can procreate. Men with power typically have more children. This was all the truer in primitive societies, where polygamy was often the norm for chieftains and kings.
- Status & Power. Equipped with Bronze weapons and horses, the Indo-Europeans would have easily subjugated the Neolithic farmers and with even greater ease Europe’s last hunter-gatherers. If they did not exterminate the indigenous men, the newcomers would have become the new ruling class, with a multitude of local kings, chieftains, and noblemen (Bronze-Age Celts and Germans lived in small village communities with a chief, each part of a small tribe headed by a king) with higher reproductive opportunities than average.
- Gender imbalance. Invading armies normally have far more men than women. Men must therefore find women in the conquered population. Wars are waged by men, and the losers suffer heavier casualties, leaving more women available to the winners.
- Aggressive warfare. The Indo-Europeans were a warlike people with a strong heroic code emphasizing courage and military prowess. Their superior technology (metal weapons, wheeled vehicles drawn by horses) and attitude to life would have allowed them to slaughter any population that did not have organized armies with metal weapons (i.e. anybody except the Middle-Eastern civilizations).
- Genetic predisposition to conceive boys. The main role of the Y-chromosome in man’s body is to create sperm. Haplogroups are determined based on mutations differentiating Y-chromosomes. Each mutation is liable to affect sperm production and sperm motility. Preliminary research has already established a link between certain haplogroups and increased or reduced sperm motility. The higher the motility, the higher the chances of conceiving a boy. It is absolutely possible that R1b could confer a bias toward more male offspring. Even a slightly higher percentage of male births would significantly contribute to the replacement of other lineages with the accumulation effect building up over a few millennia. Not all R1b subclades might have this boy bias. The bias only exist in relation to other haplogroups found in a same population. It is very possible that the fairly recent R1b subclades of Western Europe had a significant advantage compared to the older haplogroups in that region, notably haplogroup I2 and E-V13. Read more” ref
“Replacement of patrilineal lineages following this model quickly becomes exponential. Imagine 100 Indo-European men conquering a tribe of 1000 indigenous Europeans (a ratio of 1:10). War casualties have resulted in a higher proportion of women in the conquered population. Let’s say that the surviving population is composed of 700 women and 300 men. Let’s suppose that the victorious Indo-European men end up having twice as many children reaching adulthood as the men of the vanquished tribe. There is a number of reason for that. The winners would take more wives, or take concubines, or even rape women of the vanquished tribe. Their higher status would guarantee them greater wealth and therefore better nutrition for their offspring, increasing the chances of reaching adulthood and procreating themselves.” ref
“An offspring ratio of 2 to 1 for men is actually a conservative estimate, as it is totally conceivable that Bronze-Age sensibilities would have resulted in killing most of the men on the losing side, and raping their women (as attested by the Old Testament). Even so, it would only take a few generations for the winning Y-DNA lineages to become the majority. For instance, if the first generation of Indo-Europeans had two surviving sons per man, against only one per indigenous man, the number of Indo-European paternal lineages would pass to 200 individuals at the second generation, 400 at the third, 800 at the fourth, and 1600 at the fifth, and so on. During that time indigenous lineages would only stagnate at 300 individuals for each generation.” ref
“Based on such a scenario, the R1b lineages would have quickly overwhelmed the local lineages. Even if the Indo-European conquerors had only slightly more children than the local men, R1b lineages would become dominant within a few centuries. Celtic culture lasted for over 1000 years in Continental Europe before the Roman conquest putting an end to the privileges of the chieftains and nobility. This is more than enough time for R1b lineages to reach 50 to 80% of the population.” ref
“The present-day R1b frequency forms a gradient from the Atlantic fringe of Europe (highest percentage) to Central and Eastern Europe (lowest), the rises again in the Anatolian homeland. This is almost certainly because agriculture was better established in Eastern, then Central Europe, with higher densities of population, leaving R1b invaders more outnumbered than in the West. Besides, other Indo-Europeans of the Corded Ware culture (R1a) had already advanced from modern Russia and Ukraine as far west as Germany and Scandinavia. It would be difficult for R1b people to rival with their R1a cousins who shared similar technology and culture. The Pre-Celto-Germanic R1b would therefore have been forced to settled further west, first around the Alps, then overtaking the then sparsely populated Western Europe.” ref
The Balkanic and Asian branch (Z2103) corresponding to haplogroup R1b
“Haak et al. (2015) tested six Y-DNA samples from the eastern reaches of Yamna culture, in the Volga-Ural region, and all of them turned out to belong to haplogroup R1b. Four of them were positive for the Z2103 mutation. IN all likelihood, R1b-Z2103 was a major lineage of the Poltavka culture, which succeeded to the Yamna culture between the Volga River and the Ural mountains. It eventually merged with the Abashevo culture (presumably belonging chiefly to R1a-Z93) to form the Sintashta culture. Through a founder effect or through political domination, R1a-Z93 lineages would have outnumbered R1b-Z2103 after the expansion to Central and South Asia, although important pockets of Z2103 survived, notably in Bashkorostan, Turkmenistan, and Uyghurstan (Chinese Turkestan).” ref
“R1b-Z2103 would have become an Indo-Iranian lineage like R1a-Z93. This is true of two Z2103 subclades in particular: L277.1 and L584. The former is found in Russia to Central Asia then to India and the Middle East, just like the R1a-L657 subclade of Z93. It can be associated with the Andronovo culture and Bactria–Margiana Archaeological Complex, as well as the Indo-Aryan migrations. R1b-L584 is found especially in Iran, northern Iraq, the South Caucasus, and Turkey, and correlates more with the Iranian branch of Indo-Europeans, which includes Persians, Kurds, and Scythians.” ref
Anatolian branch corresponding to haplogroup R1b
“The Hittites (c. 2000-1178 BCE) were the first Indo-Europeans to defy (and defeat) the mighty Mesopotamian and Egyptian empires. There are two hypotheses regarding the origins of the Hittites. The first is that they came from the eastern Balkans and invaded Anatolia by crossing the Bosphorus. That would mean that they belonged either to the L23* or the Z2103 subclade. The other plausible scenario is that they were an offshoot of the late Maykop culture, and that they crossed the Caucasus to conquer the Hattian kingdom (perhaps after being displaced from the North Caucasus by the R1a people of the Catacomb culture). In that case, the Hittites might have belonged to the R1b-Z2103 or the R1b-PF7562 subclade. The first hypothesis has the advantage of having a single nucleus, the Balkans, as the post-Yamna expansion of all Indo-European R1b. The Maykop hypothesis, on the other hand, would explain why the Anatolian branch of IE languages (Hittite, Luwian, Lydian, Palaic) is so archaic compared to other Indo-European languages, which would have originated in Yamna rather than Maykop.” ref
“There is substantial archaeological and linguistic evidence that Troy was an Indo-European city associated with the Steppe culture and haplogroup R1b. The Trojans were Luwian speakers related to the Hittites (hence Indo-European), with attested cultural ties to the culture of the Pontic-Caspian Steppe. The first city of Troy dates back to 3000 BCE, right in the middle of the Maykop period. Troy might have been founded by Maykop people as a colony securing the trade routes between the Black Sea and the Aegean. The founding of Troy happens to coincide exactly with the time the first galleys were made. Considering the early foundation of Troy, the most likely of the two Indo-European paternal haplogroups would be R1b-M269 or L23.” ref
“The Phrygians and the Proto-Armenians are two other Indo-European tribes stemming from the Balkans. Both appear to have migrated to Anatolia around 1200 BCE, during the ‘great upheavals’ of the Eastern Mediterranean (see below). The Phrygians (or Bryges) founded a kingdom (1200-700 BCE) in west central Anatolia, taking over most of the crumbling Hittite Empire. The Armenians crossed all Anatolia until Lake Van and settled in the Armenian Highlands. Nowadays 30% of Armenian belong to haplogroup R1b, the vast majority to the L584 subclade of Z2103 (=> see The Indo-European migrations to Armenia).” ref
“Most of the R1b found in Greece today is of the Balkanic Z2103 variety. There is also a minority of Proto-Celtic S116/P312 and of Italic/Alpine Celtic S28/U152. Z2103 could have descended from Albania or Macedonia during the Dorian invasion (see below), thought to have happened in the 12th century BCE. Their language appears to have been close enough to Mycenaean Greek to be mutually intelligible and easy for locals to adopt. The Mycenaeans might have brought some R1b (probably also Z2103) to Greece, but their origins can be traced back through archaeology to the Catacomb culture and the Seima-Turbino phenomenon of the northern forest-steppe, which would make them primarily an R1a tribe.” ref
“Greek and Anatolian S116 and some S28 lineages could be attributed to the La Tène Celtic invasions of the 3rd century BCE. The Romans also certainly brought S28 lineages (=> see Genetics of the Italian people), and probably also the Venetians later on, notably on the islands. Older clades of R1b, such as P25 and V88, are only a small minority and would have come along E1b1b, G2a, and J2 from the Middle East.” ref
The great upheavals circa 1200 BCE corresponding to haplogroup R1b
“1200 BCE was a turning point in European and Near-Eastern history. In Central Europe, the Urnfield culture evolved into the Hallstatt culture, traditionally associated with the classical Celtic civilization, which was to have a crucial influence on the development of ancient Rome. In the Pontic Steppe, the Srubna culture make way to the Cimmerians, a nomadic people speaking an Iranian or Thracian language. The Iron-age Colchian culture (1200-600 BCE) starts in the North Caucasus region. Its further expansion to the south of the Caucasus corresponds to the first historical mentions of the Proto-Armenian branch of Indo-European languages (circa 1200 BCE). In the central Levant the Phoenicians start establishing themselves as significant maritime powers and building their commercial empire around the southern Mediterranean.” ref
“But the most important event of the period was incontestably the destruction of the Near-Eastern civilizations, possibly by the Sea Peoples. The great catastrophe that ravaged the whole Eastern Mediterranean from Greece to Egypt circa 1200 BCE is a subject that remains controversial. The identity of the Sea Peoples has been the object of numerous speculations. What is certain is that all the palace-based societies in the Near-East were abruptly brought to an end by tremendous acts of destruction, pillage, and razing of cities. The most common explanation is that the region was invaded by technologically advanced warriors from the north. They could have been either Indo-Europeans descended from the Steppe via the Balkans, or Caucasian people (G2a, J1, J2a, T1a) linked with the expansion of the earlier Kura-Araxes culture to eastern Anatolia and the Levant.” ref
“The Hittite capital Hattusa was destroyed in 1200 BCE, and by 1160 BCE the empire had collapsed, probably under the pressure of the Phrygians and the Armenians coming from the Balkans. The Mycenaean cities were ravaged and abandoned throughout the 12th century BCE, leading to the eventual collapse of Mycenaean civilization by 1100 BCE. The kingdom of Ugarit in Syria was annihilated and its capital never resettled. Other cities in the Levant, Cyprus, and Crete were burned and left abandoned for many generations. The Egyptians had to repel assaults from the Philistines from the East and the Libyans from the West – two tribes of supposed Indo-European origin. The Lybians were accompanied by mercenaries from northern lands (the Ekwesh, Teresh, Lukka, Sherden, and Shekelesh), whose origin is uncertain, but has been placed in Anatolia, Greece, and/or southern Italy.” ref
“The devastation of Greece followed the legendary Trojan War (1194-1187 BCE). It has been postulated that the Dorians, an Indo-European people from the Balkans (probably coming from modern Bulgaria or Macedonia), invaded a weakened Mycenaean Greece after the Trojan War, and finally settled in Greece as one of the three major ethnic groups. The Dorian regions of classical Greece, where Doric dialects were spoken, were essentially the southern and eastern Peloponnese, Crete, and Rhodes, which is also the part of Greece with the highest percentage of R1b-Z2103.” ref
“Another hypothesis is that the migration of the Illyrians from north-east Europe to the Balkans displaced previous Indo-European tribes, namely the Dorians to Greece, the Phrygians to north-western Anatolia, and the Libu to Libya (after a failed attempt to conquer the Nile Delta in Egypt). The Philistines, perhaps displaced from Anatolia, finally settled in Palestine around 1200 BCE, unable to enter Egypt.” ref
Other migrations of R1b
“Other migrations occurred from Europe to the Near East and Central Asia during the Antiquity and Middle Ages. R1b-S28 (U152) was found in Romania, Turkey, northern Bashkortostan (a staggering 71.5% of the local population according to Myres et al.), and at the border of Kazakhstan and Kyrgyzstan. Some of it was surely brought by the La Tène Celts, known to have advanced along the Danube, and created the Galatian kingdom in central Anatolia. The rest could just as well be Roman, given that R1b-S28 is the dominant form of R1b in the Italian peninsula.” ref
“Some have hypothesized that some “lost” Roman legions went as far as Central Asia or China and never came back, marrying local women and leaving their genetic marker in isolated pockets in Asia. A more prosaic version is that Roman merchants ended up in China via the Silk Road, which existed since the 2nd century BCE. A small percentage of Western European R1b subclades were also found among Christian communities in Lebanon. They are most likely descendants of the crusaders.” ref
The lactase persistence allele and R1b cattle pastoralists
“Lactose (milk sugar) is an essential component of breast milk consumed by infants. Its digestion is made possible by an enzyme, called lactase, which breaks down lactose in simple sugars that can be absorbed through the intestinal walls and into the bloodstream. In most mammals (humans included), the production of the lactase enzyme is dramatically reduced soon after weaning. As a result, older children and adults become lactose intolerant. That is true of a big part of the world population. Some people possess a genetic mutation that allows the production of lactase through adulthood. This is called lactase persistence (LP). Lactase persistence is particularly common among Northwest Europeans, descended from the ancient Celtic and Germanic people, and in parts of Africa where cattle herding has been practiced for thousands of years. The highest incidence for the lactase persistence alleles, known to geneticists as -13,910*T (rs4988235) and -22018*A (rs182549), are found among Scandinavian, Dutch, British, Irish, and Basque people. Sub-Saharan populations with lactase persistence have different mutations, such as -14010*C, -13915*G, and -13907*G.” ref
“R1b men are thought to be the first people on earth to successfully domesticate cattle and to develop a lifestyle based on cattle husbandry and herding during the Pre-Pottery Neolithic (see Neolithic section). Looking for pasture for their cows, R1b tribes migrated from the Near East to the savannah of North Africa (which has since underwent desertification and become the Sahara) and to the Pontic Steppe in southern Russia and Ukraine. For several millennia no other human population was so dependent on cattle for their survival as these R1b tribes.” ref
“It is known that most Neolithic herding societies consumed at least some animal milk and even made cheese from it (since cheese contains less lactose and is easier to digest for people who are lactose intolerant). In most of Europe, the Middle East, and South Asia, people essentially herded goats and sheep, better suited to mountainous environment of the Mediterranean basin, Anatolia, and Iran. Goats and sheep could also be kept easily inside villages by sedentary cereal cultivators, while cows needed vast pastures for grazing, which were particularly scarce in the Middle East. Domesticated cattle were sometimes found in small number among other Neolithic populations, but the ones that relied almost entirely on them were the R1b tribes of the Pontic Steppe and North Africa. To this very day, semi-nomadic pastoralists in the Sahel, such as the Fulani and the Hausa, who are descended from Neolithic R1b-V88 migrants from the Near East, still maintain primarily herds of cattle. It is among these cattle herders that selective pressure for lactase persistence would have been the strongest.” ref
“There has been speculations among geneticists and evolutionary biologists regarding the origin of the lactase persistence allele in Europeans. Over 100 ancient DNA samples have been tested from Mesolithic, Neolithic, and Bronze Age Europe and Syria, and the -13910*T allele has been found only in Late Neolithic/Chalcolithic and Bronze Age individuals. The origin of the mutation does not really matter, since it could have been present at low frequencies in the human gene pool for tens of thousands of years before it underwent postive selective pressure among cattle-herding societies. What is certain is that individuals from Bronze Age cultures associated with the arrival of Indo-European speakers from the Pontic Steppe already possessed relatively high percentages of the LP allele. For example, the LP allele was found at a frequency of 27% (see Schilz 2006) among the 13 individuals from the Lichtenstein Cave in Germany, who belonged to the Urnfield culture, and were a mix of Y-haplogroups R1b, R1a, and I2a2b.” ref
“Nowadays, the LP allele is roughly proportional to the percentage of R1b, and to a lower extent R1a, found in a population. In the British Isles, the Low Countries and south-west Scandinavia, where LP is the highest in the world, the combined percentage of R1a and R1b exceeds 70% of the population. In Iberia, the highest percentage of LP is observed among the Basques, who have the highest percentage of R1b. In Italy, LP is most common in the north, like R1b. The lowest incidence of LP in Europe are found in South Italy, Greece, and the Balkans, the regions that have the least R1b lineages.” ref
“Tishkoff et al. (2017) confirmed that the Hausa and the Fulani, two Sahel tribes with high incidence of R1b-V88, possessed the same LP allele as Europeans, but that East African pastoralist populations with a high prevalence of the lactase persistence trait possess a completely different mutation, which arose independently. This finding is the strongest evidence so far that the -13,910*T allele originated with the first R1b cattle herders in the Near East, who are the ancestors of both the Indo-Europeans and of African R1b-V88 tribes.” ref
R1 populations spread genes for light skin, blond hair, and red hair
“There is now strong evidence that both R1a and R1b people contributed to the diffusion of the A111T mutation of the SLC24A5, which explains approximately 35% of skin tone difference between Europeans and Africans, and most variations within South Asia. The distribution pattern of the A111T allele (rs1426654) of matches almost perfectly the spread of Indo-European R1a and R1b lineages around Europe, the Middle East, Central Asia, and South Asia. The mutation was probably passed on in the Early neolithic to other Near Eastern populations, which explains why Neolithic farmers in Europe already carried the A111T allele (e.g. Keller 2012 p.4, Lazaridis 2014 suppl. 7), although at lower frequency than modern Europeans and southern Central Asians.” ref
“The light skin allele is also found at a range of 15 to 30% in various ethnic groups in northern sub-Saharan Africa, mostly in the Sahel and savannah zones inhabited by tribes of R1b-V88 cattle herders like the Fulani and the Hausa. This would presuppose that the A111T allele was already present among all R1b people before the Pre-Pottery Neolithic split between V88 and P297. R1a populations have an equally high incidence of this allele as R1b populations. On the other hand, the A111T mutation was absent from the 24,000-year-old R* sample from Siberia, and is absent from most modern R2 populations in Southeast India and Southeast Asia.” ref
“Consequently, it can be safely assumed that the mutation arose among the R1* lineage during the late Upper Paleolithic, probably some time between 20,000 and 13,000 years ago. Fair hair was another physical trait associated with the Indo-Europeans. In contrast, the genes for blue eyes were already present among Mesolithic Europeans belonging to Y-haplogroup I. The genes for blond hair are more strongly correlated with the distribution of haplogroup R1a, but those for red hair have not been found in Europe before the Bronze Age, and appear to have been spread primarily by R1b people (=> see The origins of red hair).” ref
What were the original mtDNA lineages of Neolithic R1b tribes in the Near East?
“R1b tribes are thought to have domesticated cattle in that region 10,500 years ago, yet only moved across the Caucasus some time between 7,500 and 6,500 years ago. For three or four millennia, semi-nomadic R1b herders were bound to have intermingled with some of the Near Eastern or Caucasian neighbors. One way of determining what mt-haplogroups R1b tribes carried at the very beginning of the Neolithic, is to compare the above haplogroups with those of African ethnic groups known to possess elevated percentages of R1b-V88. The best studied group are the Fulani, whose mtDNA includes three European-looking haplogroups H, J1b1a, U5, and V making up about 15% of their total maternal lineages.” ref
“These haplogroups have been identified in all four Central African countries sampled, confirming a strong correlation with haplogroup R1b. However, their H, V, and U5 could have come from the Berbers of Northwest Africa. The Berbers also carry R1b-V88, but it’s possible that some of it came from different Neolithic migrations, including a re-expansion from Iberia, as Berbers carry H1, H3, V1a1a, V5, and U5b1b1, lineages that are all found in the Iberian peninsula. U5b1b1 descends from Mesolithic West Europeans, but at present, it is not yet clear how the other haplogroups reached Iberia or Northwest Africa. One hypothesis is that they came from the Near East during the Neolithic, perhaps with R1b-V88 tribes.” ref
“African R1b-V88 and Eurasian haplogroup R1b-P297 split roughly 10,000 years ago, almost certainly in Eastern Europe, where they carried mostly mt-haplogroup U5. Toward the end of the last glaciation, some R1b men would have migrated from Eastern Europe to the region of modern Kurdistan accompanied by women belonging to mtDNA U5. Soon after they arrived J1b1a (and maybe V) would have been the first indigenous Near Eastern lineages assimilated by R1b tribes. R1b-V88 might have assimilated H1 and H3 women in the Levant before moving to North Africa, but that remains highly hypothetical.” ref

ref, ref, ref, ref, ref, ref, ref, ref, ref, ref
Proto-Indo-European society
“Proto-Indo-European society is the reconstructed culture of Proto-Indo-Europeans, the ancient speakers of the Proto-Indo-European language, ancestor of all modern Indo-European languages. Archaeologist David W. Anthony and linguist Donald Ringe distinguish three different cultural stages in the evolution of the Proto-Indo-European language:
- Early (4500–4000 BCE), the common ancestor of all attested Indo-European languages, before the Anatolian split (Cernavodă culture; 4000 BCE); associated with the early Khvalynsk culture,
- Classic, or “post-Anatolian” (4000–3500 BCE), the last common ancestor of the non-Anatolian languages, including Tocharian; associated with the late Khvalynsk and Repin cultures,
- Late (3500–2500 BCE), in its dialectal period due to the spread of the Yamnaya horizon over a large area.” ref
Early Khvalynsk (4900–3900) and Proto-Indo-European society
“Domesticated cattle were introduced around 4700 BCE from the Danube valley to the Volga–Ural steppes where the Early Khvalynsk culture (4900–3900 BCE) had emerged, associated by Anthony with the Early Proto-Indo-European language. Cattle and sheep were more important in ritual sacrifices than in diet, suggesting that a new set of cults and rituals had spread eastward across the Pontic-Caspian steppes, with domesticated animals at the root of the Proto-Indo-European conception of the universe. Anthony attributes the first and progressive domestication of horses, from taming to actually working with the animal, to this period. Between 4500–4200 BCE, copper, exotic ornamental shells, and polished stone maces were exchanged across the Pontic–Caspian steppes from Varna, in the eastern Balkans, to Khvalynsk, near the Volga river. Around 4500, a minority of richly decorated single graves, partly enriched by imported copper items, began to appear in the steppes, contrasting with the remaining outfitted graves.” ref
“The Anatolian distinctive sub-family may have emerged from a first wave of Indo-European migration into southeastern Europe around 4200–4000 BCE, coinciding with the Suvorovo–to–Cernavoda I migration, in the context of a progression of the Khvalynsk culture westwards towards the Danube area, from which had also emerged the Novodanilovka (4400–3800) and Late Sredny Stog (4000–3500 BCE) cultures.” ref
Late Khvalynsk/Repin (3900–3300) and Proto-Indo-European society
“Steppe economies underwent a revolutionary change between 4200 to 3300 BCE, in a shift from a partial reliance on herding, when domesticated animals were probably used principally as a ritual currency for public sacrifices, to a later regular dietary dependence on cattle, and either sheep or goat meat and dairy products. The Late Khvalynsk and Repin cultures (3900–3300 BCE), associated with the classic (post-Anatolian) Proto-Indo-European language, showed the first traces of cereal cultivation after 4000 BCE, in the context of a slow and partial diffusion of farming from the western parts of the steppes to the east. Around 3700–3300 BCE, a second migration wave of proto-Tocharian speakers towards South Siberia led to the emergence of the Afanasievo culture (3300–2500 BCE).” ref
“The spoke-less wheeled wagon was introduced to the Pontic-Caspian steppe around 3500 BCE from the neighboring North Caucasian Maykop culture (3700–3000 BCE), with which Proto-Indo-Europeans traded wool and horses. Interactions with the hierarchical Maykop culture, itself influenced by the Mesopotamian Uruk culture, had notable social effects on the Proto-Indo-European way of life. Meanwhile, the Khvalynsk-influenced cultures that had emerged in the Danube-Donets region after the first migration gave way to the Cernavodă (4000–3200 BCE), Usatovo (3500–2500 BCE), Mikhaylovka (3600–3000 BCE) and Kemi Oba (3700—2200 BCE) cultures, from west to east respectively.” ref
Yamnaya period (3300–2600) and Proto-Indo-European society
“The Yamnaya horizon, associated with the Late Proto-Indo-European language (following both the Anatolian and Tocharian splits), originated in the Don–Volga region before spreading westwards after 3300 BCE, establishing a cultural horizon founded on kurgan funerals that stretched over a vast steppe area between the Dnieper and Volga rivers. It was initially a herding-based society, with limited crop cultivation in the eastern part of the steppes, while the Dnieper–Donets region was more influenced by the agricultural Tripolye culture. Paleolinguistics likewise postulates Proto-Indo-European speakers as a semi-nomadic and pastoral population with subsidiary agriculture.” ref
“Bronze was introduced to the Pontic-Caspian steppes during this period. Following the Yamnaya expansion, long-distance trade in metals and other valuables, such as salt in the hinterlands, probably brought prestige and power to Proto-Indo-European societies. However, the native tradition of pottery making was weakly developed. The Yamnaya funeral sacrifice of wagons, carts, sheep, cattle, and horses was likely related to a cult of ancestors requiring specific rituals and prayers, a connection between language and cult that introduced the Late Proto-Indo-European language to new speakers. Yamnaya chiefdoms had institutionalized differences in prestige and power, and their society was organized along patron-client reciprocity, a mutual exchange of gifts and favors between their patrons, the gods, and human clients. The average life expectancy was fairly high, with many individuals living to 50–60 years old. The language itself appeared as a dialect continuum during this period, meaning that neighboring dialects differed only slightly between each other, whereas distant language varieties were probably no longer mutually intelligible due to accumulated divergences over space and time.” ref
“As the steppe became dryer and colder between 3500–3000 BCE, herds needed to be moved more frequently in order to feed them sufficiently. Yamnaya distinctive identity was thus founded on mobile pastoralism, permitted by two earlier innovations: the introduction of the wheeled wagon and the domestication of the horse. Yamnaya herders likely watched over their cattle and raided on horseback, while they drove wagons for the bulk transport of water or food. Light-framework dwellings could be easily assembled and disassembled to be transported on pack animals.” ref
“Another climate change that occurred after around 3000 BCE led to a more favorable environment allowing for grassland productivity. Yamnaya new pastoral economy then experienced a third wave of rapid demographic expansion, that time towards Central and Northern Europe. Migrations of Usatovo people towards southeastern Poland, crossing through the Old European Tripolye culture from around 3300 BCE, followed by Yamnya migrations towards the Pannonian Basin between 3100–2800 BCE, are interpreted by some scholars as movements of pre-Italic, pre-Celtic and pre-Germanic speakers.” ref
“The Proto-Indo-European language probably ceased to be spoken after 2500 BCE as its various dialects had already evolved into non-mutually intelligible languages that began to spread across most of western Eurasia during the third wave of Indo-European migrations (3300–1500 BCE). Indo-Iranian languages were introduced to Central Asia, present-day Iran, and South Asia after 2000 BCE.” ref
Class Structure and Proto-Indo-European society
“It is generally agreed that Proto-Indo-European society was hierarchical, with some form of social ranking and various degrees of social status. It is unlikely however that they had a rigidly stratified structure, or castes such as are found in historical India. There was a general distinction between free persons and slaves, typically prisoners of war or debtors unable to repay a debt. The free part of society was composed of an elite class of priests, kings, and warriors, along with the commoners, with each tribe following a chief (*wiḱpots) sponsoring feasts and ceremonies, and immortalized in praise poetry.” ref
“The presence of kurgan graves prominently decorated with dress, body ornaments, and weaponry, along with well-attested roots for concepts such as “wealth” (*h₂ép-), “to be in need” (*h₁eg-) or “servant” (*h₂entbʰi-kʷolos, “one who moves about on both sides”; and *h₂upo-sth₂-i/o-, “one standing below”), indicate that a hierarchy of wealth and poverty was recognized. Some graves, larger than the average and necessitating a considerable number of people to be built, likewise suggest a higher status given to some individuals. These prestigious funerals were not necessarily reserved to the wealthiest person. Smiths in particular were given sumptuous graves, possibly due to the association of smithery with magic during the early Bronze Age. In general, such graves were mostly occupied by males in the eastern Don-Volga steppes, while they were more egalitarian in the western Dnieper-Donets region.” ref
Kinship and Proto-Indo-European society
“Linguistics has allowed for the reliable reconstruction of a large number of words relating to kinship relations. These all agree in exhibiting a patriarchal, patrilocal and patrilineal social fabric. Patrilocality is confirmed by lexical evidence, including the word *h₂u̯edh-, ‘to lead (away)’, being the word that denotes a male wedding a female. Rights, possessions, and responsibilities were consequently reckoned to the father, and wives were to reside after marriage near the husband’s family, after the payment of a bride-price.” ref
“The household (*domos) was generally ruled by the senior male of the family, the *dems-potis (‘master of the household’), and could also consist of his children, grandchildren, and perhaps unrelated slaves or servants. His wife probably also played a complementary role: some evidence suggest that she would have kept her position as the mistress (*pot-n-ih₂) of the household in the event her husband dies, while the eldest son would have become the new master. The Proto-Indo-European expansionist kinship system was likely supported by both marital exogamy (the inclusion of foreign women through marriage) and the exchange of foster children with other families and clans, as suggested by genetic evidence and later attestations from Indo-European-speaking groups.” ref
“Once established, the family lasted as long as the male stock of its founder endured, and clan or tribal founders were often portrayed as mythical beings stemming from a legendary past in Indo-European traditions. In this form of kinship organization, the individual’s genetic distance from the clan’s founding ancestor determined his social status. But if he was of exceptional prowess or virtue, the same individual could in his turn gain social prestige among the community and eventually found his own descent-group.” ref
“In the reconstructed lexicon linking the individual to the clan, *h₂erós means a ‘member of one’s own group’, ‘one who belongs to the community’ (in contrast to an outsider). It gave way to the Indo-Iranian *árya (an endonym), and probably to the Celtic *aryos (‘noble, freeman’), the Hittite arā- (‘peer, companion’), and the Germanic *arjaz (‘noble, distinguished’). It is unlikely however that the term had an ethnic connotation, and we do not know if Proto-Indo-European speakers had a term to designate themselves as a group. Another word, *h₁leudhos, means ‘people’, ‘freemen’ in a more general way.” ref
Patron-client and Proto-Indo-European society
“Proto-Indo-European had several words for ‘leader’: *tagós was a general term derived from *tā̆g- (‘set in place, arrange’); *h₃rḗǵs meant a ruler who also had religious functions, with the Roman rex sacrorum (‘king of the sacred’) as a heritage of the priestly function of the king; *w(n̩)nákts designated a ‘lord’ and possessed a feminine equivalent, *wnáktih₂ (a ‘queen’); while the *wiḱpots (or *wiḱ-potis) was the chief of the settlement (*weiḱs), the seat of a tribe, clan or family.” ref
“Public feasts sponsored by such patrons were a way for them to promote and secure a political hierarchy built on the unequal mobilization of labor and resources, by displaying their generosity towards the rest of the community. Rivals competed publicly through the size and complexity of their feasts, and alliances were confirmed by gift-giving and promises made during those public gatherings. The host of the feast was called the *ghosti-potis, the ‘lord of the guests’, who honored the immortal gods and his mortal guests with gifts of food, drink, and poetry.” ref
Guest-Host and Proto-Indo-European society
“Vertical social inequalities were partly balanced by horizontal mutual obligations of hospitality between guests and hosts. According to Anthony, the domestication of horses and the introduction of the wagon in the Pontic-Caspian steppe between 4500 to 3500 BCE led to an increase in mobility across the Yamnaya horizon, and eventually to the emergence of a guest-host political structure. As various herding clans began to move across the steppes, especially during harsh seasons, it became necessary to regulate local migrations on the territories of tribes that had likely restricted these obligations to their kins or co-residents (*h₂erós) until then. In Proto-Indo-European, the term *ghós-ti-, whose original meaning must have been “table companion”, could either mean a host or a guest. The connotation of an obligatory reciprocity between both guests and hosts has persisted in descendant cognates, such as Latin hospēs (“foreigner, guest; host”), Old English ġiest (“stranger, guest”), or Old Church Slavonic gostĭ (“guest”) and gospodĭ (“master”).” ref
“Guests and hosts were indeed involved in a mutual and reciprocal relationship bound by oaths and sacrifices. The giving and receiving of favors was accompanied by a set of ritual actions that indebted the guest to show hospitality to his host at any time in the future. The obligation could even be heritable: Homer’s warriors, Glaukos and Diomedes, stopped fighting and presented gifts to each other when they learned that their grandfathers had shared a guest-host relationship. Violations of the guest-host obligations were considered immoral, illegal, and unholy: in Irish law, refusing hospitality was deemed a crime as serious as murder. The killing of a guest was also greeted with a singular revulsion, as was the abuse of hospitality.” ref
Legal System and Proto-Indo-European society
“Because of the archaic nature of traditional legal phraseology—which preserves old forms and meaning for words—and the necessity for legal sentences to be uttered precisely the same way each time to remain binding, it is possible to securely reconstruct some elements of the Proto-Indo-European legal system. For instance, the word *serk– (‘to make a circle, complete’) designated a type of compensation where the father (or master) had to either pay for the damages caused by his son (or slave), or surrender the perpetrator to the offended party. It is attested by a common legal and linguistic origin in both Roman and Hittite laws. Another root denoting a compensation, *kwey-, had the meanings of ‘blood-price‘, ‘vengeance’ or ‘guilt’ in daughter languages, suggesting that it was specifically applied to the restitution for theft or violence.” ref
“Law was apparently designed to preserve the ‘order’ (*h₂értus) of the universe, with the underlying idea that the cosmic harmony should be maintained, be it in the physical universe or the social world. There was however probably no public enforcement of justice, nor were there formal courts as we know them today. Contractual obligations were protected by private individuals acting as sureties: they pledged to be responsible for payments of debts incurred by someone else if the latter defaulted. In case of litigation, one could either take matter into their own hands, for instance by barring someone from accessing their property to compel payment, or bring the case before judges (perhaps kings) that included witnesses. The word for ‘oath’, *óitos, derives from the verb *h₁ei- (‘to go’), after the practice of walking between slaughtered animals as part of taking an oath.” ref
“The root *h₂értus (from *h₂er-, ‘to fit’) is associated with the concept of a cosmic order, that is which is ‘fitting, right, ordered’. It is one of most securely reconstructed Proto-Indo-European words, with cognates attested in most sub-families: Latin artus (‘joint’); Middle High German art (‘innate feature, nature, fashion’); Greek artús (ἀρτύς, ‘arrangement’), possibly arete (ἀρετή, ‘excellence’); Armenian ard (արդ, ‘ornament, shape’); Avestan arəta- (‘order’) and ṛtá (‘truth’); Sanskrit ṛtú- (ऋतु, ‘right time, order, rule’); Hittite āra (????????????, ‘right, proper’); Tocharian A ārt- (‘to praise, be pleased with’).” ref
Trifunctional Hypothesis and Proto-Indo-European society
“The trifunctional hypothesis, proposed by Georges Dumézil, postulates a tripartite ideology reflected in a threefold division between a clerical class (encompassing both the religious and social functions of the priests and rulers), a warrior class (connected with the concepts of violence and braveness), and a class of farmers or husbandmen (associated with fertility and craftsmanship), on the basis that many historically-known groups speaking Indo-European languages show such a division. Dumézil initially contended that it derived from an actual division in Indo-European societies, but later toned down his approach by representing the system as functions or general organizing principles. Dumézil’s theory has been influential and some scholars continue to operate under its framework, although it has also been criticized as aprioristic and too inclusive, and thus impossible to be proved or disproved.” ref
Rituals and Proto-Indo-European society
Proto-Indo-Europeans practiced a polytheistic religion centered on sacrificial rites of cattle and horses, probably administered by a class of priests or shamans. Animals were slaughtered (*gʷʰn̥tós) and dedicated to the gods (*déiwos) in the hope of winning their favor. The king as the high priest would have been the central figure in establishing good relations with the other world. The Khvalynsk culture, associated with early Proto-Indo-European, had already shown archeological evidence for the sacrifice of domesticated animals. Proto-Indo-Europeans also had a sacred tradition of horse sacrifice for the renewal of kinship involving the ritual mating of a queen or king with a horse, which was then sacrificed and cut up for distribution to the other participants in the ritual.” ref
“Although we know little about the role of magic in Proto-Indo-European society, there is no doubt that it existed as a social phenomenon, as several branches attest the use of similarly worded charms and curses, such as ones against worms. Furthermore, incantations and spells were frequently regarded as one of the three categories of medicine, along with the use of surgical instruments and herbs or drugs. Since the earliest evidence for the burning of the plant was found in Romanian kurgans dated 3,500 BCE, some scholars suggest that cannabis was first used as a psychoactive drug by Proto-Indo-Europeans during ritual ceremonies, a custom they eventually spread throughout western Eurasia during their migrations. Descendant cognates of the root *kanna- (“cannabis”) have been proposed in Sanskrit śaná, Greek kánnabis (κάνναβις), Germanic *hanipa (German Hanf, English hemp), Russian konopljá, Albanian kanëp, Armenian kanap and Old Prussian knapios. Other linguists suggest that the common linguistic inheritance does not date back to the Indo-European period and contend that the word cannabis likely spread later across Eurasia as a Wanderwort (‘wandering word’), ultimately borrowed into Ancient Greek and Sanskrit from a non-Indo-European language.” ref
Poetry and Proto-Indo-European society
“Poetry and songs were central to Proto-Indo-European society. The poet-singer was the society’s highest-paid professional, possibly a member of a hereditary profession that ran in certain families, the art passing from father to son as the poet had to acquire all the technical aspects of the art and to master an extensive body of traditional subject matter. He performed against handsome rewards—such as gifts of horses, cattle, wagons, and women—and was held in high esteem. In some cases, the poet-singer had a stable relationship with a particular noble prince or family. In other cases, he traveled about with his dependants, attaching himself to one court after another.” ref
“A transmitter of inherited cultural knowledge, the poet sang as a recall of the old heroic times, entrusted with telling the praises of heroes, kings, and gods. Composing sacred hymns ensured the gods would in turn bestow favorable fate to the community, and for kings that their memory would live on many generations. A lexeme for a special song, the *erkw (“praise of the gift”) has been identified in early Proto-Indo-European. Such praise poems proclaimed the generosity of the gods (or a patron) and enumerated their gifts, expanding the patron’s fame, the path to immortality otherwise only attainable for mortals through conspicuous acts of war or piety.” ref
“The concept of fame (*ḱléwos) was central to Proto-Indo-European poetry and culture. Many poetic dictions built on this term can be reconstituted, including *ḱléwos wéru (“wide fame”), *ḱléwos meǵh₂ (“great fame”), *ḱléuesh₂ h₂nróm (“the famous deeds of men, heroes”), or *dus-ḱlewes (“having bad repute”). Indo-European poetic tradition was probably oral-formulaic: stock formulas, such as the imperishable fame (*ḱléwos ń̥dʰgʷʰitom), the swift horses (*h₁ōḱéwes h₁éḱwōs), the eternal life (*h₂iu-gʷih₃), the metaphor of the wheel of the sun (*sh₂uens kʷekʷlos), or the epithet man-killer (*hₐnr̥-gʷhen), attached to Hektor and Rudra alike, were transmitted among poet-singers to fill out traditional verse-lines in epic song lyrics. The task of the Indo-European poet was to preserve over the generations the famous deeds of heroes. He would compose and retell poems based on old and sometimes obscure formulations, reconnecting the motifs with his own skills and improvisations. Poetry was therefore associated with the acts of weaving words (*wékʷos webh-) and crafting speech (*wékʷos teḱs-).” ref
Warfare: Kóryos and Proto-Indo-European society
“Although Proto-Indo-Europeans have been often cast as warlike conquerors, their reconstructed arsenal is not particularly extensive. There is no doubt that they possessed archery, as several words with the meaning of “spear” (*gʷéru ; *ḱúh₁los), “pointed stick” (*h₂eiḱsmo), or “throwing spear” (*ǵʰai-só-s) are attested. The term *wēben meant a “cutting weapon”, probably a knife, and *h₂/₃n̩sis a “large offensive knife”, likely similar to bronze daggers found across Eurasia around 3300–3000 BCE. Proto-Indo-Europeans certainly did not know swords, which appeared later around 2000–1500 BCE. The ax was known as *h₄edʰés, while the word *spelo/eh₂ designated a wooden or leather shield. The term *leh₂wós meant “military unit” or “military action”, while *teutéh₂- might have referred to the “adult male with possession” who would mobilize during warfare, perhaps originally a Proto-Indo-European term meaning “the people under arms.” ref
“A number of scholars propose that Proto-Indo-European rituals included the requirement that young unmarried men initiate into manhood by joining a warrior-band named *kóryos. They were led by a senior male and lived off the country by hunting and engaging in raiding and pillaging foreign communities. Kóryos members served in such brotherhoods (Männerbunden) for a number of years before returning home to adopt more respectable identities as mature men. During their initiation period, the young males wore the skin and bore the names of wild animals, especially wolves (*wl̩kʷo) and dogs (*ḱwōn), in order to assume their nature and escape the rules and taboos of their host society.” ref
“Most kurgan stelae found in Pontic-Caspian steppe feature a man wearing with a belt and weapons carved on the stone. In later Indo-European traditions, notably the (half-)naked warrior figures of Germanic and Celtic art, *kóryos raiders wore a belt that bound them to their leader and the gods, and little else. The tradition of kurgan stelae featuring warriors with a belt is also common in Scythian cultures. A continuity of an “animal-shaped raid culture” has been also postulated based on various elements attested in later Indo-European-speaking cultures, such as the Germanic Berserkers, the Italic Ver Sacrum, and the Spartan Crypteia, as well as in the mythical Celtic fianna and Vedic Maruts, and in the legend of the werewolf (“man-wolf”), found in Greek, Germanic, Baltic and Slavic traditions alike.” ref
“In a mostly patriarchal economy based on bride competition, the escalation of the bride-price in periods of climate change could have resulted in an increase in cattle raiding by unmarried men. Scholars also suggest that, alongside the attractiveness of the patron-client and the guest-host relationships, the *kóryos could have played a key role in diffusing Indo-European languages across most of Eurasia.” ref
Personal names and Proto-Indo-European society
“The use of two-word compound words for personal names, typically but not always ascribing some noble or heroic feat to their bearer, is so common in Indo-European languages that it is certainly an inherited feature. These names often belonged in early dialects to the class of compound words that in the Sanskrit tradition are called bahuvrihi. As in Vedic bahuvrihi (literally “much-rice”, meaning “one who has much rice”), those compounds are formed as active structures indicating possession and do not require a verbal root. From the Proto-Indo-European personal name *Ḱléwos-wésu (lit. “good-fame”, meaning “possessing good fame”) derive the Liburnian Vescleves, the Greek Eukleḗs (Εὐκλεής), the Old Persian Huçavah, the Avestan Haosravah-, and the Sanskrit Suśráva.” ref
“A second type of compound consists of a noun followed by a verbal root or stem, describing an individual performing an action. Compounds more similar to synthetics are found in the Sanskrit Trasá-dasyus (“one who causes enemies to tremble”), the Greek Archelaus (Ἀρχέλαος, “one who rules people”), and the Old Persian Xšayāršan (“one who rules men”).” ref
“Many Indo-European personal names are associated with the horse (*h₁éḱwos) in particular, which expressed both the wealth and nobility of their bearer, including the Avestan Hwaspa (“owning good horses”), the Greek Hippónikos (“winning by his horses”), or the Gaulish Epomeduos (“master of horses”). Since domestic animals also served to sacrifice, there were often used as exocentric structures in compound names (the bearers are not ‘horses’ themselves but ‘users of horses’ in some way), in contrast to endocentric personal names rather associated with wild animals like the wolf, for instance in the German Adolf (“a noble wolf”) or the Serbian Dobrovuk (“a good wolf”).” ref
Economy and Proto-Indo-European society
“Proto-Indo-Europeans possessed a Neolithic mixed economy based on livestock and subsidiary agriculture, with a wide range of economic regimes and various degrees of mobility that could be expected across the large Pontic-Caspian steppe. Tribes were typically more influenced by farming in the western Dnieper–Donets region, where cereal cultivation was practiced, while the eastern Don–Volga steppes were inhabited by semi-nomadic and pastoral populations mostly relying on herding. Proto-Indo-European distinguished between unmovable and movable wealth (*péḱu, the “livestock”). As for the rest of society, the economy was founded on reciprocity. A gift always entailed a counter-gift, and each party was bound to the other in a mutual relationship cemented by trust.” ref
Trade and Proto-Indo-European society
“The early Khvalynsk culture, located in the Volga–Ural steppes and associated with early Proto-Indo-European, had trade relationship with Old European cultures. Domesticated cattle, sheep, and goats, as well as copper, were introduced eastward from the Danube valley around 4700–4500 BCE. Copper objects show an artistic influence from Old Europe, and the appearance of sacrificed animals suggests that a new set of rituals emerged following the introduction of herding from the west. The Old European Tripolye culture continued to influence the western part of the steppes, in the Dnieper–Donets region, where the Yamnaya culture was more agricultural and less male-centered.” ref
“Proto-Indo-European speakers also had indirect contacts with Uruk around 3700–3500 through the North Caucasian Maikop culture, a trade route that introduced the wheeled wagon into the Caspian-Pontic steppes. Wheel-made pottery imported from Mesopotamia were found in the Northern Caucasus, and Maikop chieftain was buried wearing Mesopotamian symbols of power—the lion paired with the bull. The late Khvalynsk and Repin cultures probably traded wool and domesticated horses in exchange, as suggested by the widespread appearance of horses in archeological sites across Transcaucasia after 3300 BCE. Socio-cultural interactions with Northwest Caucasians have been proposed, on the ground that the Proto-Indo-European language shows a number of lexical parallels with Proto-Northwest Caucasian. Proto-Indo-European also exhibits lexical loans to or from other Caucasian languages, particularly Proto-Kartvelian.” ref
“Proto-Indo-European probably also had trade relationships with Proto-Uralic speakers around the Ural Mountains. Words for “sell” and “wash” were borrowed in Proto-Uralic, and words for “price” and “draw, lead” were introduced in the Proto-Finno-Ugric language. James P. Mallory suggested that the expansion of the Uralic languages across the northern forest zone might have been stimulated by organizational changes within Uralic forager societies, resulting partly from interaction with more complex, hierarchical Proto-Indo-European and (later) Indo-Iranian pastoral societies at the steppe/forest-steppe ecological border.” ref
Technology and Proto-Indo-European society
“From the reconstructable lexicon, it is clear that Proto-Indo-Europeans were familiar with wheeled vehicles—certainly horse-drawn wagons (*weǵʰnos)—as they knew the wheel (*kʷekʷlóm), the axle (*h₂eḱs-), the shaft (*h₂/₃éih₁os), and the yoke (*yugóm). Although wheels were most likely not invented by Proto-Indo-Europeans, the word *kʷekʷlóm is a native derivation of the root *kʷel– (“to turn”) rather than a borrowing, suggesting short contacts with the people who introduced the concept to them.” ref
“The technology used was a solid wheel made of three planks joined together with their outer edges trimmed to a circle. The swift chariot with spoked wheels, which made the mode of transport much more rapid and lighter, appeared later within the Sintashta culture (2100–1800 BCE), associated with the Indo-Iranians. As the word for “boat” (*néh₂us) is widely attested across the language groups, the means of transport (likely a dugout canoe) was certainly known by Proto-Indo-Europeans.” ref
“The vocabulary associated with metallurgy is very restricted and at best we can attest the existence of copper/bronze, gold, and silver. The basic word for “metal” (*h₂ey-es) is generally presumed to mean “copper” or a copper-tin alloy of “bronze”. “Gold” is reliably reconstructed as *h₂eusom, and *h₂erǵ-n̩t-om designated a “white metal” or “silver”. Proto-Indo-Europeans were also familiar with the sickle (*sr̩po/eh₂), the awl (*h₁óleh₂) for working leather or drilling wood, and used a primitive plough (*h₂érh₃ye/o) made of a curved and forked branch.” ref
“The term for “oven” or “cooking vessel” (*h₂/₃ukʷ) has been reconstructed based on four branches, as for “baking” (*bʰōg-) and “boiling” (*yes-). They certainly drank beer (*h₂elut) and mead (*médʰu), and the word for “wine” (*wóinom) has been proposed, although this remains a debated issue. Proto-Indo-Europeans produced textile, as attested by the reconstructed roots for wool (*wĺh₂neh₂), flax (*linom), sewing (*syuh₁-), spinning (*(s)pen-), weaving (*h₂/₃webʰ-) and plaiting (*pleḱ-), as well as needle (*skʷēis) and thread (*pe/oth₂mo). They were also familiar with combs (*kes) and ointments with salve (*h₃engʷ-).” ref
Animals and Proto-Indo-European society
“Animals (mammals in particular) are fairly abundant in the reconstructed lexicon. We can ascribe about seventy-five names to various animal species, but it hardly recovers all the animals to have been distinguished in the proto-language. While *kʷetwor-pod designated a four-footed animal (tetrapod), *gʷyéh₃wyom seems to have been the general term for animals, derived from the root *gʷyeh₃-, “to live”. Proto-Indo-European speakers also made a distinction between wild animals (*ǵʰwḗr) and the livestock (*péḱu).” ref
Domesticated Animals and Proto-Indo-European society
“The reconstructed lexicon suggests a Neolithic economy with extensive references to domesticated animals. They were familiar with cows (*gʷṓus), sheep (*h₃ówis), goats (*díks, or *h₂eiĝs), and pigs (*sūs ; also *pórḱos, “piglet”). They knew dogs (*ḱwōn), milk (*ǵl̩ákt; also *h₂melǵ-, “to milk”) and dairy foods, wool (*wĺh₂neh₂) and woolen textiles, agriculture, wagons, and honey (*mélit). The domestication of the horse (*h₁éḱwos), thought to be an extinct Tarpan species, probably originated with these peoples, and scholars invoke this innovation as a factor contributing to their increased mobility and rapid expansion.” ref
“The dog was perceived as a symbol of death and depicted as the guardian of the Otherworld in Indo-European cultures (Greek Cerberus, Indic Śarvarā, Norse Garmr). The mytheme possibly stems from an older Ancient North Eurasian belief, as evidenced by similar motifs in Native American and Siberian mythology, in which case it might be one of the oldest mythemes recoverable through comparative mythology. In various Indo-European traditions, the worst throw at the game of dice was named the “dog”, and the best throw was known as the “dog-killer”. Canine teeth of dogs were frequently worn as pendants in Yamnaya graves in the western Pontic steppes, particularly in the Ingul valley.” ref
Wild Animals and Proto-Indo-European society
“Linguistic evidence suggests that Proto-Indo-European speakers were also in contact with various wild animals, such as red foxes (*wl(o)p), wolves (*wl̩kʷo), bears (*h₂ŕ̩tḱos), red deers (*h₁elh₁ēn), elks (moose) (*h₁ólḱis), eagles (*h₃or), otters (*udrós), snakes (*h₁ógʷʰis), mice (*mūs ; from *mus-, “to steal”), or trouts (*lóḱs). Some of them were featured in mythological and folkloric motifs. Goats draw the chariots of the Norse and Indic gods Thor and Pushan, and they are associated with the Baltic god Perkūnas and the Greek god Pan. The words for both the wolf and the bear underwent taboo deformation in a number of branches, suggesting that they were feared as symbols of death in Proto-Indo-European culture.” ref
“In Indo-European culture, the term “wolf” is generally applied to brigands and outlaws who live in the wild. Ritual and mythological concepts connected with wolves, in some cases similar with Native American beliefs, may represent a common Ancient North Eurasian heritage: mai-coh meant both “wolf” and “witch” among Navajos, and shunk manita tanka a “doglike powerful spirit” among Sioux, while the Proto-Indo-European root *ṷeid (“knowledge, clairvoyance”) designated the wolf in both Hittite (ṷetna) and Old Norse (witnir), and a “werewolf” in Slavic languages (Serb vjedo-gonja, Slovenian vedanec, Ukrainian viščun).” ref
Beliefs: Proto-Indo-European mythology
“The reconstructed cosmology of the proto-Indo-Europeans shows that the ritual sacrifice of cattle, cows in particular, was at the root of their beliefs, as the primordial condition of the world order. The myth of *Trito, the first warrior, involves the liberation of cattle stolen by a three-headed serpent named *Ngwhi. After recovering the wealth of the people, Trito eventually offered the cattle to the priest in order to ensure the continuity of the cycle of giving between gods and humans. The creation myth could have rationalized raiding as the recovery of cattle that the gods had intended for the people who sacrificed properly. Many Indo-European cultures preserved the tradition of cattle raiding, which they often associated with epic myths. Georges Dumézil suggested that the religious function was represented by a duality, one reflecting the magico-religious nature of the priesthood, while the other is involved in religious sanction to human society (especially contracts), a theory supported by common features in Iranian, Roman, Scandinavian and Celtic traditions. The study of astronomy was not much developed among Proto-Indo-Europeans, and they probably had established names for only a few individual stars and star-groups (e.g. Sirius, Ursa Major).” ref
“The basic word for “god” in proto-Indo-European is *deiwós (“celestial”), itself a derivative of *dei– (“to shine, be bright”). On the other hand, the word for “earth” (*dʰéǵʰōm) is at root of both “earthly” and “human”, as it is notably attested in the Latin cognates humus and homo. This suggests a hierarchical conception of the status of mankind regarding the gods, confirmed by the use of the term “mortal” (*mr̩tós) as a synonym of “human” as opposed to the never-dying gods in Indo-European traditions. The idea is expressed in the Homeric phrase “of the immortal gods and of men who walk on earth”.” ref
Proto-Indo-European beliefs were influenced by a resistant animistic substratum, and the few names that can be reconstructed based upon both linguistic (cognates) and thematic (reflexes) evidence are the cosmic and elemental deities: the ‘Daylight-Sky’ (*Dyḗus), his partner ‘Earth’ (*Dʰéǵʰōm), his daughter the ‘Dawn’ (*H₂éwsōs), and his Twin Sons, the ‘Sun’ (*Séh₂ul) and the Sun-Maiden, and deities of winds, waters, fire, rivers and springs. The Proto-Indo-European creation myth tells of a primordial sacrifice performed by the first man *Manu (“Man”) on his twin brother *Yemo (“Twin”), from whom emerged the cosmological elements. Other deities, such as the weather-god *Perkʷunos and the guardian of roads and herds, *Péh₂usōn, are probably late innovations since they are attested in a restricted number of traditions, Western (European) and Graeco-Aryan, respectively.” ref


The “wheel” related word list
“Most linguists argue that the PIEs (Proto-Indo-Europeans) did have words for wheel. The candidates put forward for wheel or wagon-related words are nine reconstructed PIE word forms. These are:
- *hurki , argued to mean “wheel”
- *roteh2, argued to mean “wheel”
- *kwékwlo-, argued to mean “wheel”
- *kwelh1-, argued to mean “turn” perhaps in the sense of a turning wheel.
- *h2eks-, argued to mean “axle”
- *h2ih3s-, argued to mean “thill” or “wagon shaft”
- *wéĝh-, argued to mean “convey in a vehicle”
- *h3nebh-, argued to mean “nave” or “wheel hub”
- *iugó-, argued to mean “yoke” ref
Proto-Indo-European language
“Proto-Indo-European (PIE) is the reconstructed common ancestor of the Indo-European language family. Its proposed features have been derived by linguistic reconstruction from documented Indo-European languages. No direct record of Proto-Indo-European exists. Far more work has gone into reconstructing PIE than any other proto-language, and it is the best understood of all proto-languages of its age. The majority of linguistic work during the 19th century was devoted to the reconstruction of PIE or its daughter languages, and many of the modern techniques of linguistic reconstruction (such as the comparative method) were developed as a result.” ref
“PIE is hypothesized to have been spoken as a single language from 4500 to 2500 BCE during the Late Neolithic to Early Bronze Age, though estimates vary by more than a thousand years. According to the prevailing Kurgan hypothesis, the original homeland of the Proto-Indo-Europeans may have been in the Pontic–Caspian steppe of eastern Europe. The linguistic reconstruction of PIE has provided insight into the pastoral culture and patriarchal religion of its speakers.” ref
“As speakers of Proto-Indo-European became isolated from each other through the Indo-European migrations, the regional dialects of Proto-Indo-European spoken by the various groups diverged, as each dialect underwent shifts in pronunciation (the Indo-European sound laws), morphology, and vocabulary. Over many centuries, these dialects transformed into the known ancient Indo-European languages. From there, further linguistic divergence led to the evolution of their current descendants, the modern Indo-European languages. Today, the descendant languages of PIE with the most native speakers are Spanish, English, Portuguese, Hindustani (Hindi and Urdu), Bengali, Russian, Punjabi, German, Persian, French, Marathi, Italian, and Gujarati.” ref
“PIE is believed to have had an elaborate system of morphology that included inflectional suffixes (analogous to English child, child’s, children, children’s) as well as ablaut (vowel alterations, as preserved in English sing, sang, sung, song) and accent. PIE nominals and pronouns had a complex system of declension, and verbs similarly had a complex system of conjugation. The PIE phonology, particles, numerals, and copula are also well-reconstructed. Asterisks are used as a conventional mark of reconstructed words, such as *wódr̥, *ḱwṓ, or *tréyes; these forms are the reconstructed ancestors of the modern English words water, hound, and three, respectively.” ref
“Commonly proposed subgroups of Indo-European languages include Italo-Celtic, Graeco-Aryan, Graeco-Armenian, Graeco-Phrygian, Daco-Thracian, and Thraco-Illyrian. There are numerous lexical similarities between the Proto-Indo-European and Proto-Kartvelian languages due to early language contact, though some morphological similarities—notably the Indo-European ablaut, which is remarkably similar to the root ablaut system reconstructible for Proto-Kartvelian—may suggest a higher-level phylogenetic relationship.” ref
“The Lusitanian language was a marginally attested language spoken in areas near the border between present-day Portugal and Spain. The Venetic and Liburnian languages known from the North Adriatic region are sometimes classified as Italic. Albanian and Greek are the only surviving Indo-European descendants of a Paleo-Balkan language area, named for their occurrence in or in the vicinity of the Balkan peninsula. Most of the other languages of this area—including Illyrian, Thracian, and Dacian—do not appear to be members of any other subfamilies of PIE, but are so poorly attested that proper classification of them is not possible. Forming an exception, Phrygian is sufficiently well-attested to allow proposals of a particularly close affiliation with Greek, and a Graeco-Phrygian branch of Indo-European is becoming increasingly accepted.” ref
List of Indo-European languages
“The Indo-European languages include some 449 (SIL estimate, 2018 edition) language families spoken by about or more than 3.5 billion people (roughly half of the world population). Most of the major languages belonging to language branches and groups of Europe, and western and southern Asia, belong to the Indo-European language family. Therefore, Indo-European is the biggest language family in the world by number of mother-tongue speakers (but not by number of languages in which it is the 3rd or 5th biggest). Eight of the top ten biggest languages, by number of native speakers, are Indo-European. One of these languages, English, is the de facto World Lingua Franca with an estimate of over one billion second-language speakers.” ref
“Each subfamily or linguistic branch in this list contains many subgroups and individual languages. Indo-European language family has 10 known branches or subfamilies, of which eight are living and two are extinct. The relation of Indo-European branches, how they are related to one another and branched from the ancestral proto-language is a matter of further research and not yet well known. There are some individual Indo-European languages that are unclassified within the language family, they are not yet classified in a branch and could be members of their own branch. The 449 Indo-European languages identified in the SIL estimate, 2018 edition, are mostly living languages, however, if all the known extinct Indo-European languages are added, they number more than 800 or close to one thousand. This list includes all known Indo-European languages, living and extinct.” ref
“A distinction between a language and a dialect is not clear-cut and simple because there is, in many cases, several dialect continuums, transitional dialects, and languages and also because there is no consensual standard to what amount of vocabulary, grammar, pronunciation, and prosody differences there is a language or there is a dialect. (Mutual intelligibility can be a standard but there are closely related languages that are also mutual intelligible to some degree, even if it is an asymmetric intelligibility.) Because of this, in this list, several dialect groups and some individual dialects of languages are shown (in italics), especially if a language is or was spoken by a large number of people and over a big land area, but also if it has or had divergent dialects. The contact between different peoples and languages, especially as a result of European colonization, also gave origin to the many pidgins, creoles, and mixed languages that are mainly based in Indo-European languages (many of which are spoken in island groups and coastal regions).” ref
The ancestral population and language, Proto-Indo-Europeans that spoke Proto-Indo-European, estimated to have lived about 4500 BCE (around 6500 years ago), at some time in the past, starting about 4000 BCE (around 6000 years ago) expanded through migration and cultural influence. This started a complex process of population blend or population replacement, acculturation, and language change of peoples in many regions of western and southern Eurasia. This process gave origin to many languages and branches of this language family. At the end of the second millennium BCE Indo-European speakers were many millions and lived in a vast geographical area in most of western and southern Eurasia (including western Central Asia). In the following two millennia, the number of speakers of Indo-European languages increased even further.” ref
“By geographical area, Indo-European languages remained spoken in big land areas, although most of western Central Asia and Asia Minor was lost to another language family (mainly Turkic) due to Turkic expansion, conquests, and settlement (after the middle of the first millennium AD and the beginning and middle of the second millennium CE respectively) and also to Mongol invasions and conquests (that changed Central Asia ethnolinguistic composition). Another land area lost to non-Indo-European languages was today’s Hungary due to Magyar/Hungarian (Uralic language speakers) conquest and settlement. However, in the second half of the second millennium CE, Indo-European languages expanded their territories to North Asia (Siberia), through Russian expansion, and North America, South America, Australia, and New Zealand as the result of the age of European discoveries and European conquests through the expansions of the Portuguese, Spanish, French, English and the Dutch. (These peoples had the biggest continental or maritime empires in the world and their countries were major powers.)” ref
Michelsberg culture
“The Michelsberg culture (German: Michelsberger Kultur (MK)) is an important Neolithic culture in Central Europe. Its dates are c. 4400–3500 BCE. Its conventional name is derived from that of an important excavated site on Michelsberg (short for Michaelsberg) hill near Untergrombach, between Karlsruhe and Heidelberg (Baden-Württemberg), Germany. The Michelsberg culture belongs to the Central European Late Neolithic. Its distribution covered much of West Central Europe, along both sides of the Rhine. A detailed chronology, based on pottery, was produced in the 1960s by the German archaeologist Jens Lüning.” ref
“The Michelsberg culture emerges in northeastern France c. 4400 BCE. Genetic evidence suggests that it originated through a migration of peoples from the Paris Basin. Its people appear to trace their origins to Mediterranean farmers expanding from the southwest and of the Linear Pottery culture of Central Europe. Shortly after its emergence in northeastern France, the Michelsberg culture expands rapidly throughout central Germany, northeastern France, eastern Belgium, and the southwestern Netherlands. These areas had previously been occupied by cultures derived from the Linear Pottery culture (LBK), with whom the Michelsberg culture shares surprisingly little cultural or genetic affinity.” ref
“Archaeological evidence suggests that the Michelsberg expansion was accompanied by violence. The Michelsberg culture has strong affinities to the Chasséen culture of central France. Archaeological evidence strongly suggests that colonists from the Michelsberg culture played an instrumental role in establishing the Funnelbeaker culture of Northern Europe, which brought agriculture to southern Scandinavia. The Michelsberg culture also displays close affinities to the cultures of the Neolithic British Isles. The spread of agriculture into the British Isles by colonists from the continent happens at almost exactly the same time as in Scandinavia, suggesting that the two events are connected. The Michelsberg culture ends about c. 3500 BCE. It is succeeded in its core area by the Wartberg culture, with which it displays strong signs of continuity.” ref
“Since 2008 the fortified settlement of Kapellenberg near Hofheim and Frankfurt (one of the largest of the Michelsberg culture) has been investigated by the Römisch-Germanisches Zentralmuseum, Leibniz Archaeological Research Institute. The settlement has been described as representing “the beginnings of urbanism,” already in 4000 BCE. The overall site was 45 hectares in size with an internal settlement covering 26 hectares, containing numerous rectangular houses and surrounded by a rampart. A large tumulus (burial mound) was built at the center of the settlement between 4200–4100 BCE, indicating the influence of the Castellic culture in Brittany, where giant burial mounds containing megalithic tombs (such as Tumiac and Saint-Michel) were built c. 4500 BCE for elite males described by some researchers as ‘divine kings’.” ref
“The Castellic mounds contained large quantities of jade axes (the jade originally imported from the Italian Alps), as well jewelry made from callaïs (variscite and turquoise) imported from south-western Spain. Jade axes have similarly been found at the Kappellenberg, attesting to an exchange network of prestige goods associated with elites as well as the trade in salt. The Kappellenberg tumulus and jade axes indicate that “a socio-political hierarchisation process linked to the emergence of high-ranking elites” was underway in the Rhine valley at the same time as similar developments were occurring in Brittany (Castellic culture) and the Paris basin (Cerny culture). The settlement at Schierstein might have housed up to several thousand inhabitants. Research so far tends to characterize MK as a culture that avoided or rejected the use of copper, but occasional finds, e.g. at Heilbronn-Klingenberg, do indicate use of that metal.” ref
“There are several tumuli, mounds of earth built up over a grave. In this area, they generally feature a passage leading to a central chamber that once held Neolithic artifacts. Three exceptionally large burial mounds are known from the Carnac and Morbihan area, dating from the mid-5th millennium BCE and known collectively as ‘Carnacéen tumuli’: Saint-Michel, Tumiac and Mané-er-Hroëk. Each of these tumuli contained a megalithic burial chamber, containing the burial of only one individual, along with numerous large polished stone axeheads, stone arm-rings, and jewelry made from callaïs (variscite and turqouise). Scientific analyses have shown that many of the axeheads are made of jade from the Italian Alps, whilst the callaïs was imported from south-western Iberia.” ref
“Archaeological evidence indicates that the callaïs was brought from Iberia by boat, across the Bay of Biscay, rather than along the coast or overland. Some of the Carnacéen jade axeheads are up to 46 cm in length and may have taken over a thousand hours to produce, on top of the time required to quarry the material and transport it to Carnac. The extraordinary nature of these burials, the scale of the tumuli, the distant exchange networks and effort involved, all indicate that these were the burials of extremely important elite individuals, that some researchers have described as “divine kings.” ref
“The large-scale effort and organization involved in the construction of megalithic monuments, such as the 20.6 metre-tall Grand Menhir of Er Grah, further suggests the existence of rulers or kings in the Carnac and Morbihan region. A similar situation has been described for the later megalithic culture in Ireland, which shows some close similarities to the megalithic culture in Brittany. Based on archaeological, DNA, and ethnographic evidence, it has been suggested that an elite male buried in the Newgrange passage grave, c. 3200 BCE, may have been a “god-king” and part of a “dynastic elite.” Similarities have also been noted with the Michelsberg culture in northeastern France and Germany (c. 4200 BCE), which featured large tumulus burials within fortified settlements and the use of Alpine jade axes, all associated with the emergence of “high-ranking elites.” ref
“Engravings on megalithic monuments in Carnac also feature numerous depictions of objects interpreted as symbols of authority and power, such as curved throwing weapons, axes, and sceptres. The Carnacéen tumuli were contemporary with the rich burials of the Varna culture in Bulgaria (c. 4500 BCE), which contained more gold than all excavated sites from the rest of the world combined in the 5th millennium BCE. The Varna burials attest to the existence of powerful elites in southeastern Europe at the same time as they appear in Carnac. Grave 43 of the Varna necropolis, which contained a single male buried with a scepter and numerous gold ornaments, also contained an Alpine jade axehead.” ref
“Michelsberg pottery is characterized by undecorated pointy-based tulip beakers. Finds of barley and emmer indicate an agricultural economy. Animal husbandry is indicated by bones of domesticated cattle, pig, sheep and goat. Domestic dogs have also been identified. Bones of deer and fox suggest that the MK diet was supplemented by hunting. There was no indication of a destruction of the site; nor were there any finds suggesting humans meeting a violent end. Some pits contained the remains of food stores.” ref
“Thus, the abandonment of the site may have had environmental reasons. A common suggestion is the drying up of the Rhine’s arms which used to flow by the bottom of the hill, due to an extensive dry period. As the result of such a change in climate, the area would not have easily supported agriculture any more, forcing human communities (and their livestock) to relocate. Prehistoric settlement patterns in Central Europe are generally quite volatile. The abandonment of a settlement may be part of a broader economic and social system. Thus, the Bruchsal area appears to contain several earthworks from different phases within MK.” ref
“Formal Michelsberg burials have only been recognized rarely. There is no indication of organized burial grounds, as known from the earlier Linear pottery (LBK) and Rössen cultures. Human skeletal remains, frequently disarticulated, have been found inside pits and ditches in many MK earthworks and have had considerable influence on the interpretation of such structures. Their discussion is closely connected with that of similar remains in the ditches of British Causewayed enclosures. “The MK settlement of Aue yielded eight pit graves, six containing a single individual and two containing several. The age profile of those buried is very striking, as it is limited to children under the age of seven and adults over 50 (a considerable age in Neolithic Europe).” ref
In other words, humans of the ages that must have dominated the active social and economic life of the settlement are absent. It has been suggested that their bodies may not have received formal burial, but were disposed of by excarnation, in which case the skeletal remains from rubbish pits may be the result of such activity. The same may apply to human bones found in the fills of enclosure ditches around MK settlements. It has also been suggested (hypothetically) that partially articulated remains found in such ditches may indicate that graves were placed on the surfaces adjacent to them and later washed into the ditches due to erosion. Occasionally, earthwork ditches contain more structured deposits of human bone, e.g. adult skeletons surrounded by those of children.” ref
“Such burials are probably connected to the realms of cult or ritual, as are specific depositions of offerings in some of the ditches, especially at the settlements of Aue and Scheelkopf. Here, ditches contained carefully placed complete vessels, well-preserved quern-stones, and the horns of aurochs. The latter had been neatly separated from the skulls, perhaps reflecting a special symbolic significance ascribed to that animal. A hitherto unknown aspect to MK burial practice is suggested by the recent discovery of MK burials in the Blätterhof cave near Hagen, Westphalia. Here, a full age profile appears to be represented. An unusual burial was found at Rosheim (Bas-Rhin, France). Here, the fill of a pit contained the crouched remains of an adult woman, her legs leaning against a quernstone. She appeared to have been laid onto a carefully placed packing of clay lumps, mixed with pottery and bones. Her death had been caused by some blunt impact on her skull.” ref
“Beau et al. 2017 examined the remains 22 Michelsberg people buried at Gougenheim, France. The 21 samples of mtDNA extracted belonged to the maternal haplogroups H (7 samples), K (4 samples), J (2 samples), W (1 samples), N (1 sample), U (3 samples) and T (2 samples). The examined individuals displayed genetic links to earlier farming populations of the Paris Basin, and were genetically very different from previous post-LBK cultures of the region, suggesting that the Michelsberg culture emerged through a migration of people from west. They displayed genetic links to other farmers of Western Europe, and carried substantial amounts of hunter-gatherer ancestry. The authors of the study proposed that migrations of people associated with the Michelsberg culture may have been responsible for the resurgence of hunter-gatherer ancestry observed in Central Europe during the Middle Neolithic.” ref
“Lipson et al. 2017 examined the remains of 4 individuals buried c. 4000–3000 BC at the Blätterhöhle site in modern-day Germany, ascribed to the Michelsberg culture and its successor, the Wartberg culture. The 3 samples of Y-DNA extracted belonged to the paternal haplogroups R1b1, R1 and I2a1, while the 4 samples of mtDNA extracted belonged to the maternal haplogroups U5b2a2, J1c1b1, H5 and U5b2b2. The individuals carried the high amount of about 40–50% Western Hunter-Gatherer (WHG) ancestry, with one individual displaying as much as c. 75%. Brunel et al. 2020 examined the remains of 18 individuals ascribed to the Michelsberg culture. The 2 samples of Y-DNA belonged to the paternal haplogroup I, while the 16 samples of mtDNA extracted belonged to types of the maternal haplogroups H (3 samples), K (9 samples), X (1 sample), T (2 samples) and U (1 sample).” ref
Funnelbeaker culture
“The Funnel(-neck-)beaker culture, in short TRB or TBK (German: Trichter(-rand-)becherkultur, Dutch: Trechterbekercultuur; Danish: Tragtbægerkultur; c. 4300–2800 BCE), was an archaeological culture in north-central Europe. It developed as a technological merger of local neolithic and mesolithic techno-complexes between the lower Elbe and middle Vistula rivers. These predecessors were the (Danubian) Lengyel-influenced Stroke-ornamented ware culture (STK) groups/Late Lengyel and Baden-Boleráz in the southeast, Rössen groups in the southwest and the Ertebølle-Ellerbek groups in the north. The TRB introduced farming and husbandry as major food sources to the pottery-using hunter-gatherers north of this line. The Funnelbeaker culture is named for its characteristic ceramics, beakers and amphorae with funnel-shaped tops, which were found in dolmen burials.” ref
“The TRB techno-complex is divided into a northern group including modern northern Germany and southern Scandinavia (TRB-N, roughly the area that previously belonged to the Ertebølle-Ellerbek complex), a western group in the Netherlands between the Zuiderzee and lower Elbe that originated in the Swifterbant culture, an eastern group centered on the Vistula catchment, roughly ranging from Oder to Bug, and south-central groups (TRB-MES, Altmark) around the middle and upper Elbe and Saale. Especially in the southern and eastern groups, local sequences of variants emerged. In the late 4th millennium BCE, the Globular Amphora culture (GAC) replaced most of the eastern and subsequently also the southern TRB groups, reducing the TRB area to modern northern Germany and southern Scandinavia. The younger TRB in these areas was superseded by the Single Grave culture (EGK) at about 2800 BCE. The north-central European megaliths were built primarily during the TRB era.” ref
“The Funnelbeaker culture emerged in northern modern-day Germany c. 4100 BCE. Archaeological evidence strongly suggests that it originated through a migration of colonists from the Michelsberg culture of Central Europe. The Michelsberg culture is archaeologically and genetically strongly differentiated from the preceding post-Linear Pottery cultures of Central Europe, being distinguished by increased levels of hunter-gatherer ancestry. Its people were probably descended from farmers migrating into Central Europe out of Iberia and modern-day France, who in turn were descended from farmers of the Cardial Ware cultures who had migrated westwards from the Balkans along the Mediterranean coast. Connections between the Funnelbeakers and these farmers of the Atlantic coast is supported by genetic evidence. After its establishment, the Funnelbeaker culture rapidly spread into southern Scandinavia and Poland, in what appears to have been a well-organized colonizing venture.” ref
“In southern Scandinavia it replaced the Ertebølle culture, which had maintained a Mesolithic lifestyle for about 1500 years after farming arrived in Central Europe. The emergence of the Neolithic British Isles through maritime colonization by Michelsberg-related groups occurred almost at the same time as the expansion of the Funnelbeaker culture into Scandinavia, suggesting that these events may be connected. Although they were largely of Early European Farmer (EEF) descent, people of the Funnelbeaker culture had a relatively high amount of hunter-gatherer admixture, particularly in Scandinavia, suggesting that hunter-gatherer populations were partially incorporated into it during its expansion into this region. People of the Funnelbeaker culture often had between 30% and 50% hunter-gatherer ancestry depending on the region.” ref
“During later phases of the Neolithic, the Funnelbeaker culture re-expanded out of Scandinavia southwards into Central Europe, establishing several regional varieties. This expansion appears to have been accompanied by significant human migration. The southward expansion of the Funnelbeaker culture was accompanied by a substantial increase in hunter-gatherer lineages in Central Europe. The Funnelbeaker communities in Central Europe which emerged were probably quite genetically and ethnically mixed, and archaeological evidence suggests that they were relatively violent. From the middle of the 4th millennium BCE, the Funnelbeaker culture was gradually replaced by the Globular Amphora culture on its southeastern fringes, and began to decline in Scandinavia.” ref
“In the early 3rd millennium BCE, the Corded Ware culture appeared in Northern Europe. Its peoples were of marked steppe-related ancestry and traced their origins in cultures further east. This period is distinguished by the construction of numerous defensive palisades in Funnelbeaker territory, which may be a sign of violent conflict between the Funnelbeakers, Corded Ware, and Pitted Ware. By 2650 BCE, the Funnelbeaker culture had been replaced by the Corded Ware culture. Genetic studies suggest that Funnelbeaker women were incorporated into the Corded Ware culture through intermixing with incoming Corded Ware males, and that people of the Corded Ware culture continued to use Funnelbeaker megaliths as burial grounds. Subsequent cultures of Late Neolithic, Bronze Age, and Iron Age Central Europe display strong maternal genetic affinity with the Funnelbeaker culture.” ref
“The Funnelbeaker culture was dominated by animal husbandry of sheep, cattle, pigs, and goats, but there was also hunting and fishing. Primitive wheat and barley was grown on small patches that were fast depleted, due to which the population frequently moved small distances. There was also mining (in the Malmö region) and collection of flintstone (Świętokrzyskie Mountains), which was traded into regions lacking the stone, such as the Scandinavian hinterland. The culture used copper from Silesia, especially daggers and axes. The Funnelbeaker Culture preserves the oldest dated evidence of wheeled vehicles in middle Europe.” ref
“One example is the engraving on a ceramic tureen from Bronocice in Poland on the northern edge of the Beskidy Mountains (northern Carpathian ring), which is indirectly dated to the time span from 3636 to 3373 BCE and is the oldest evidence for covered carriages in Central Europe. They were drawn by cattle, presumably oxen whose remains were found with the pot. Today it is housed in the Archaeological Museum of Cracow (Muzeum Archeologiczne w Krakowie), Poland. At Flintbek in northern Germany cart tracks dating from c. 3400 BCE were discovered underneath a megalithic long barrow. This is the earliest known direct evidence for wheeled vehicles in the world (i.e. not models or images).” ref
“Houses were centered on a monumental grave, a symbol of social cohesion. Burial practices were varied depending on region and changed over time. Inhumation seems to have been the rule. The oldest graves consisted of wooden chambered cairns inside long barrows, but were later made in the form of passage graves and dolmens. Originally, the structures were probably covered with a mound of earth and the entrance was blocked by a stone. The Funnelbeaker culture marks the appearance of megalithic tombs at the coasts of the Baltic and of the North sea, an example of which are the Sieben Steinhäuser in northern Germany.” ref
“The megalithic structures of Ireland, France and Portugal are somewhat older and have been connected to earlier archeological cultures of those areas. At graves, the people sacrificed ceramic vessels that contained food along with amber jewelry and flint-axes. Genetic analysis of several dozen individuals found in the Funnelbeaker passage grave Frälsegården in Sweden suggests that these burials were based on a patrilineal social organisation, with the vast majority of males being ultimately descended from a single male ancestor while the women were mostly unrelated who presumably married into the family. Flint-axes and vessels were also deposited in streams and lakes near the farmlands, and virtually all of Sweden’s 10,000 flint axes that have been found from this culture were probably sacrificed in water. They also constructed large cult centers surrounded by pales, earthworks, and moats. The largest one is found at Sarup on Fyn. It comprises 85,000 m2 and is estimated to have taken 8000 workdays. Another cult center at Stävie near Lund comprises 30,000 m2.” ref
“In the context of the Kurgan hypothesis (or steppe hypothesis), the culture is seen as non-Indo-European, representing a culture of Neolithic origin, as opposed to the Indo-European-language-speaking peoples (see Yamna culture) who later intruded from the east. Marija Gimbutas postulated that the political relationship between the aboriginal and intrusive cultures resulted in quick and smooth cultural morphosis into the Corded Ware culture. In the past, a number of other archaeologists proposed that the Corded Ware culture was a purely local development of the Funnelbeaker culture, but genetic evidence has since demonstrated that this was not the case.” ref
“Ancient DNA analysis has found the people who produced the Funnelbeaker culture to be genetically different from earlier hunter-gather inhabitants of the region, and are instead closely related to other European Neolithic farmers, who ultimately traced most of their ancestry from early farmers in Anatolia, with some admixture from European hunter-gatherer groups. Genetic analysis suggests that there was some minor gene flow between the producers of the Funnelbeaker culture and those of the hunter-gatherer Pitted Ware culture (which descended from earlier Scandinavian hunter-gather groups) to the north. A total of 62 males from sites attributed to the Funnelbeaker culture in Scandinavia and Germany have been sequenced for ancient DNA. Most belonged to haplogroup I2 while a smaller number belonged to R1b-V88, Q-FTF30, and G2a. MtDNA haplogroups included U, H, T, R and K.” ref
Baalberge group
“The Baalberge Group (German: Baalberger Kultur, also Baalberge-Kultur) was a late neolithic “culture” in Central Germany and Bohemia between 4000 and 3150 BCE. Because of issues with the archaeological use of the term culture it is now often referred to as the Baalberge Ceramic style (Baalberger Keramikstil). It is named after its first findspot: on the Schneiderberg at Baalberge, Salzlandkreis, Saxony-Anhalt. The Baalberge group is generally seen as part of the Funnelbeaker culture. In the Middle Elbe/Saale region it is part of Funnelbeaker phase TRB-MES II and III. An early example of the Funnelbeaker culture, the Baalberge ceramic style dates between 3800 – 3400 BCE and belongs to the central German funnelbeaker phases TRB-MES II (3800-3500 BCE) and TRB-MES III (3500-3350 BCE). It developed out of phase TRB-MES I (4100-3800 BCE), innovating under the influence of western influences (Michelsberg culture and the southeastern, late Lengyel culture). A more complex society developed after 3350 BCE in the TRB-MES IV phase, with distinct decorative styles (Salzmünd group and Walternienburg-Bernburg Culture).” ref
“The Baalberge culture was first identified as a distinct group on the grounds of pottery types by Nils Niklasson and Paul Kupka. Before this it had been included in the Bernburg type. Kupka grouped the finds belonging to the Baalberge culture together under the name “Central German Stilthouse Pottery” (mitteldeutsche Pfahlbaukeramik). Paul Grimm followed this with the first division of the material into Early, High, Late and Pre-Unetice periods in 1937. Paul Kupka and C.J. Becker put the Baalberg group in parallel with the northern Funnelbeaker culture. Joachim Preuß divided the Baalberg culture into an older and later phase using burial practices and pottery typologies. Scientific data indicates however that the divisions do not indicate chronological differences. According to Johannes Müller they instead show different social groups.” ref
“Marija Gimbutas and her followers argue that the Baalberg culture was an intrusive hybrid culture deriving ultimately from the Eurasian steppe, part of the Kurgan hypothesis. In that case it would have been an Indo-European-speaking culture. Some aspects of Baalberge burials might support this theory, such as the presence of pottery allegedly influenced by the Baden culture (an Indo-Europeanised culture according to Gimbutas) and the Bodrogkeresztúr culture and the posture of the corpses, laid on their right hand side with their legs pulled up – a posture typical of the “Yamna culture.” But other aspects of the burials are very different from burials in the east, such as the placement of the hands over the mouth in an eating gesture (which is unknown in authentic kurgan sites) and the much less marked use of red ochre. In particular, there are no signs of the steppe kurgans that characterise the Kurgan culture. Finally, comparative anatomy suggests the deceased came from a locally derived population, not from the east. Mallory therefore considers the Baalberge group a local development.” ref
“The main distribution area is the central Elbe-Saale region. Further finds occur in Mecklenburg-Vorpommern and Brandenburg. A distinctive variant is also known from Bohemia, of which numerous individual finds extend as far as Lower Austria. The distribution area extends further to the north than that of the Rössen culture. The whole settlement area is self-contained. Settlements are only barely known. Usually, they contain waste pits with characteristic material (clay, stone, bone material), hearths, and the postholes of a few individual houses in rather extended settlements (Braunsdorf, Merseburg). Houses are rectangular or square and of medium size. Pit-houses are rectangular or oval. Other pits were used for storage, waste disposal and sacrifices. A large settlement in Pirkau, Hohenmölsen was found in emergency excavations but provides no evidence of houses. There were also finds at the settlement built at Dölauer Heide in Halle which is surrounded by an embankment.” ref
“The Baalberge group is largely made up of undecorated wares with clearly defined neck, shoulder, and lower parts. They have a very round profile, varying between egg-shaped, biconical, and bulbous, with clearly differentiated bases. Incised and stamped patterns occur around the neck and shoulder. The pottery types, generally used as funerary items, were very well represented at Dölauer Heide in Halle. A grey-brown leatherlike finish is typical of the Baalberge culture. When broken, the sherds reveal a dark grey to black fabric. The complete ceramic inventory forms a self-contained type region in the usage area of the Baalberg culture. The metal finds associated with the Baalberge culture are among the oldest in the central German Neolithic. The economy was unspecialized agriculture and pastoralism. Crops included emmer, einkorn, dwarf wheat, and barley. Animals included cattle, pigs, sheep, and goats.” ref
“The graves are mostly individual. A large graveyard was found at Zauschwitz, Borna. Double graves and pit graves are also known (e.g. Schalkenburg, Quenstedt). Unusual forms include settlement burials and partial burials. In addition to simple earth graves, innovative forms with architectural elements also appear. The first tumulus graves in central Germany also come from this culture. Baalberge is the first culture in which megalithic influences in the form of grave complexes, tumulus enclosures, and cists can be detected. At the same time, heavy cits sunk in the ground or raised above it and slab graves are rare. Other grave elements can be detected, such as stone packing, wooden fittings and the combination of stone and wooden components.” ref
“Tumulus graves contain earth and stone cist graves as primary burials. In the gravemound at Latdorf in Bernburg, a narrow stone cist was found which was surrounded by a 25 metre long trapezoidal barrow. The earthen graves of the Baalberge culture usually contain inhumations, with the bodies nearly always laid out on a west-east orientation flexed on their right side. Some inhumations were enclosed in square or trapezoidal ditches. In 1966, J. Preuß recorded 116 grave-complexes in the central German habitation area. The grave complex at Stemmem, excavated by W. Matthias in 1952, is 16.4 m long and squared at both ends and was the first enclosure grave recognized as a Baalberge burial.” ref
“In 1983, G. Möbes published a number of new finds from Thuringia. In Großbrembach in Sömmerda a nearly square complex with rounded corners measuring 10.8 x 10.4 metres contains two crouching inhumations lying on their right sides and oriented south-north. The grave ditch was described as partially flat, partially trough shaped. Bright bands in the soil filling seem to indicate the inflow of water. Unetice stone packed graves around it indicate that the flat barrow must have remained well known. A similar complex was investigated in 1974 at Sommerberg near Großfahner in the proximity of Erfurt. It was a 19/17 x 15.5/14.5 metre trapezoid containing a 2.3 metre wide grave. The west side was overlapped by a small burial hut of the Corded Ware culture with several skeletons. This Corded Ware grave in the centre also shows that the main tomb must have remained well known even 1000 years after its construction. This reuse by later cultures, including the Globular Amphora culture and the Unetice culture, is common. Pottery was found as grave goods, including combinations of pitchers and cups.” ref
“A belief in an afterlife (perhaps in the grave) is suggested by the grave goods. Cultic finds include charred remains of human and animal skeletons in a pit at Melchendorf in Erfurt. At Wansleben in Eilsleben an upright human skull between two sandstone plates and below a plate covered in cattle horns attests to the practice of skull deposition. Mathieson et al. (2015) found two males carrying the paternal haplogroups I2 and R1b1b=R-PF6323, V88. Besides this, other teams only found many mtDNA examples between 3900 and 3150 BCE.” ref
Frequencies of prehistoric mtDNA and Y-DNA from the European Paleolithic to the Iron Age
Paleolithic Y-chromosomal haplogroups by chronological period
- Proto-Aurignacian (47,000 to 43,000 years ago; Eastern Europe): F
- Aurignacian Culture (43,000 to 28,000 years ago; all ice-free Europe): CT, C1a, C1b, I
- Gravettian Culture (31,000 to 24,000 years ago; all ice-free Europe): BT, CT, F, C1a2
- Solutrean Culture (22,000 to 17,000 years ago; France, Spain):?
- Epiravettian Culture (22,000 to 8,000 years ago; Italy): R1b1a
- Magdalenian Culture 17,000 to 12,000 years ago; Western Europe): IJK, I
- Epipaleolithic France (13,000 to 10,000 years ago): I
- Azilian Culture (12,000 to 9,000 years ago; Western Europe): I2 ref
Paleolithic mitochondrial haplogroups by chronological period
- Proto-Aurignacian (47,000 to 43,000 years ago; Eastern Europe): N, R*
- Aurignacian Culture (43,000 to 28,000 years ago; all ice-free Europe): M, U, U2, U6
- Gravettian Culture (31,000 to 24,000 years ago; all ice-free Europe): M, U, U2’3’4’7’8’9, U2 (x5), U5 (x5), U8c (x2)
- Solutrean Culture (22,000 to 17,000 years ago; France, Spain): U
- Epiravettian Culture (22,000 to 8,000 years ago; Italy): U2’3’4’7’8’9, U5b2b (x2)
- Magdalenian Culture 17,000 to 12,000 years ago; Western Europe): R0, R1b, U2’3’4’7’8’9, U5b (x2), U8a (x5)
- Epipaleolithic France (13,000 to 10,000 years ago): U5b1, U5b2a, U5b2b (x2)
- Epipaleolithic Germany (13,000 to 11,000 years ago): U5b1 (x2)
- Azilian Culture (12,000 to 9,000 years ago; Western Europe): U5b1h ref
Mesolithic Y-chromosomal haplogroups by country
- Ireland: I2a1b, I2a2
- Britain: IJK, I2a2 (x2)
- France: I (x3), I2a1b2
- Luxembourg: I2a1b
- Germany: I2a2a
- Spain: C1a2
- Italy: I, I2a2
- Sweden: I2a1 (x2), I2a1a1a*, I2a1b (x2), I2c2
- Estonia: R1a-YP1272
- Latvia: I2a1 (x2), I2a1b, I2a2a1, I2a2a1b (x2), Q1a2, R1b1a1a-P297 (x7)
- Lithuania: I2a1b, I2a1a2a1a-L233
- Serbia: I, I2 (x2), I2a1, I2a2, I2a2a-M223, I2a2a-Z161 (x2), R, R1b1a-L794 (x8)
- Romania: R, R1, R1b
- Ukraine: IJ, I (x4), I2, I2a1, I2a2, I2a2a, I2a2a1b1-L702 (x2), R1a, R1b1a-L794
- Russia: J, R1a1* (x3), R1a1-YP1301, R1b1a, R1b1a1a-P297 ref
Mesolithic mitochondrial haplogroups by country
Note that the very late Mesolithic Pitted Ware culture (c. 3200–2300 BCE) in Sweden is listed separately, as it is possible that intermarriages with Neolithic or Chalcolithic neighbors took place. ref
- Croatia: U5b2a5
- France: U5a2 (x2), U5b1, U5b1b
- Germany, Luxembourg: U2e, U4, U5a, U5a2c (x2), U5a2c3, U5b (x2), U5b1a, U5b1d1 (x2), U5b2a2, U5b2c1
- Greece: K1c (x2)
- Italy: U5b1
- Lithuania: U4, U5b (x3)
- Poland: U5a, U5b (x2), U5b1b
- Spain: U5b, U5b1, U5b2c1 (x2)
- Russia: C, C1g, C5d, D, H, U2e, U4 (x3), U4a, U4a1, U5a (x3), U5a1 (x2), U5a1d, T, Z1a (x2)
- Sweden: U2e1 (x2), U4b1, U5a1 (x3), U5a2, U5a2d (x2)
- Sweden (Pitted Ware): H, H1f, HV0 (x2), K1a, K1a1 (x3), T2b (x2), U, U4 (x8), U4a1, U4d (x3), U5a, U5a1a’g (x2), U5b (x2), U5b1, U5b2b1a ref
Neolithic Y-chromosomal haplogroups by culture
- Neolithic Greece (c. 9,000 to 5,200 years ago): G2a2a1b
- Starčevo–Kőrös–Criş Culture (c. 8,000 to 6,500 years ago; Southeast Europe): F (x2), G2, G2a (x5), G2a2b (x2), H2, I, I2a, I2a1
- Linear Pottery Culture (aka LBK, c. 8,000 to 6,500 years ago; Central Europe)
- Early Neolithic France (c. 7,200 to 6,700 years ago): C1a2 (x1), C1a2b (x1), G2a (x1), H2 (x2), I2a1a2 (x4), I2a1b (x1), I2a1b1a1b (x2) => Rivollat (2020) & Brunel (2020)
- Early Neolithic Baden-Württenberg (c. 7,300 to 6,700 years ago): CT (x2), C1a2b (x1), F (x2), H2 (x10), I2a2a1b (x1)=> Rivollat (2020)
- Early Neolithic Saxony-Anhalt (c. 7,250 to 6,850 years ago): F (x2), G2 (x1), G2a2a (x7), G2a2b (x3), G2a2b2a1 (x2), T1a (x2) => Brandt (2013), Mathieson (2015) & Rivollat (2020)
- Early Neolithic Austria (c. 7,500 to 6,500 years ago): C1a2 (x1), G2a2b2a3 (x1), J2 (x1) => Mathieson (2017)
- Early Neolithic Hungary (c. 7,300 to 6,900 years ago): C1a2 (x1), F (x1), G2a2a1 (x1), G2a2b (x2), G2a2b2a (x1), G2a2b2b1 (x1), I1 (x1) => Szécsényi-Nagy (2015) & Lipson (2017)
- Sopot & Lengyel Cultures (7,000 to 5,400years ago; Central Europe): E1b1b-M78, I2a, J2
- Cucuteni-Trypillian Culture (c. 7,500 to 4,750 years ago; Romania, Moldova, western Ukraine): E, G2a, G2a2b2a (x2), G2a2b2a1a1b1a1a1
- Cardium Pottery Culture (c. 8,400 to 4,700 years ago; Mediterranean Europe): E1b1b-V13, G2a (x3), I2a1b1, R1b1c-V88
- Middle Neolithic France (c. 6,700 to 5,300 years ago): E1b1a1a1a1c2c (x1), E1b1b (x1), E1b1b1a1 (x3), G2a2a (x4), G2a2b2a (x12), H2 (x7), IJK (x1), I (x3), I1a2b4 (x1), I2 (x2), I2a (x1), I2a1a (x11), I2a1b (x5), I2a2b (x2) => Rivollat (2020) & Brunel (2020)
- Late Neolithic France (c. 5,500 to 3,200 years ago): G2a (x1), G2a1a1a1a1a (x1), G2a2b2b (x1), I2 (x1) => Brunel (2020)
- Atlantic Megalithic Culture (c. 7,000 to 4,000 years ago; Western Europe):
- Megalithic Ireland (c. 6,700 to 5,350 years ago): H2a (x2), I (x4), I2a1a2 (x4), I2a1a2-S21825 (x7), I2a2 (x4), I2a2a-L1195 (x21) => Sanchez-Quinto (2019) & Cassidy et al. (2020)
- Megalithic Britain (c. 7,700 to 4,550 years ago): I (x1), I2 (x1), I2a1b (x13), I2a2 (x6) => Olalde (2018) & Sanchez-Quinto (2019)
- Neolithic Britain (c. 5,900 to 4,450 years ago): I (x2), I2a1b (x1), I2a2 (x1), I2a2a (x11) => Brace (2019)
- Megalithic Sweden (c. 5,300 to 4,600 years ago): I2a1b (x4) => Sanchez-Quinto (2019)
- Megalithic France (c. 4,800 to 4,800 years ago): I2a1 (x2) => Lacan (2011)
- Megalithic Iberia (c. 5,900 to 5,350 years ago): H2 (x1), I2a1a (x1), I2a2 (x1) => Mathieson (2015) & Martiniano (2017)
- Funnelbeaker Culture (aka TRB, c. 6,000 to 4,700 years ago; Northern Europe):
- Baalberge group (c. 5,800 to 5,350 years ago; central Germany): I, R1
- Salzmünde group (5,400 to 5,000 years ago: East Germany): G2a2a (x2), I2a1b1a (x2) ref
Neolithic mitochondrial haplogroups by culture
- Neolithic Greece (c. 9,000 to 5,200 years ago): X2b
- Cardium Pottery Culture* (c. 8,400 to 4,700 years ago; Mediterranean Europe): H (x3), H1 (x3), H3 (x5), H4a1a (x2), HV0 (x2), J1 (x6), J1c3, K(x2), K1a (x3), K1a2a (x2), K1a4a1, N, N1a1a1, T2b (x4), T2c1d, U, U5 (x5), U5b1c, V (x2), X1, X2 (x4), X2c
- Starčevo–Kőrös–Criş Culture (c. 8,000 to 6,500 years ago; Southeast Europe): H, H5, HV0, J, J1c (x4), K (x4), K1 (x2), K1a (x7), N1a1, N1a1a, N1a1a1, R3, T1a, T2, T2b (x7), T2c, T2e, U3, U4, V (x2), V6, W (x2), X2 (x3)
- Linear Pottery Culture (aka LBK, c. 8,000 to 6,500 years ago; Central Europe): H (x12), H1, H1j, H5 (x2), H26b, HV (x2), J (x7), J1c17, K (x10), K1a (x8), K1a2, K1a3a3, K2a5, N1a1a (x3), N1a1a1, N1a1a1a, N1a1a1a1, N1a1a1a2, N1a1a1a3 (x5), N1a1a3, T (x3), T1a, T2 (x3), T2b (x9), T2b23 (x2), T2b23a, T2c (x2), T2c1, T2c1b, T2e (x4), U2, U3, U5a1, U5a1a’g, U5b, U5b2c, V, W (x2), X2d1
- Alföld Linear Pottery Culture (c. 7,850 to 7,350 years ago: Hungary): H, J1c1, N1a
- Rubané récent du Bassin parisien (aka RRBP, c. 7,500 to 6,300 years ago: northern France): H (x5), H1 (x8), H3 (x2), J (x3), J1, J2, K (x8), N1a (x3), T (x2), U, U4, U5 (x5), U5b, U5b2b1a, V (x2), X, X2b
- Neolithic Alsace (c. 7,500 to 6,300 years ago: northern France): H, H1, HV, J1 (x2), K, K1a, K1a4a1e, N1a1a1a (x2), U5, V, X
- Rössen Culture (c. 6,600 to 6,300 years ago; Germany, Low Countries, France, Switzerland): H1, H5, H5b, H16, H89, HV0 (x2), K (x2), N1a1a, T2e, T2f7, U5b, V, X2c
- Schöningen group (c. 6,200 to 5,950 years ago; Germany): H (x2), H1e7, H10i, HV, J, J1c, J2b1a, K (x3), K1a (x3), N1a1a1a3, T2b, T2c, T2f, U5b3, U5b2a2c, U8b1b, W1c (x2), X2b1’2’3’4’5’6
- Other samples from Middle Neolithic Germany (c. 5,900 to 5,000 years ago): H1c3, H2 (x2), H5 (x3), H11a, HV0 (x2), J, T2b, U3a1, U5b2a2 (x2), U5b2a5, X2
- Sopot & Lengyel Cultures (7000 to 5400 years ago; Central Europe): H (x2), H1, H5, H39, HV0, J1c1, N1a1a1a, T2b (x3), U5a, U8b1b
- Cucuteni-Trypillian Culture (c. 7,500 to 4,750 years ago; Romania, Moldova, western Ukraine): R0, H(x3), H5*, H5b*, H1b1*, HV, HV0, HV6-24, J, T2a1b1, T2b, U8b1a2b, U8b1b
- Atlantic Megalithic Culture (c. 7,000 to 4,000 years ago; Western Europe): H1, H3, HV0, J, K (x2), K1a, K1a1, K1a1b1 (x2), N1a, T2a1b, T2b (x2), U4, U5b (x3), U5b1, U5b2b3, U5b3, V, X (x2), X2
- Neolithic Britain (c. 6,000 to 4,500 years ago): H (x10), HV (x1), J (x6), K1 (x16), T2 (x5), U2 (x1), U5a (x3), U5b (x4), U8 (x1), V (x1), X2 (x2), W (x1)
- Funnelbeaker Culture (aka TRB, c. 6,000 to 4,700 years ago; Northern Europe): H (x3), H1, H24, J1d5, J2b1a, K1a5, T2b
- Baalberge group (c. 5,800 to 5,350 ybp ; central-east Germany): H (x3), H1e1a, H7d5, HV, J, K1a (x2), N1a1a, T1a1, T2b, T2c (x2), T2e1, U5b2a2, U8a1a, X, X2c
- Walternienburg-Bernburg group (c. 5,100 to 4,700 years ago; central-east Germany): H, H1e1a3, H5, K1, K1a (x2), T2b, U5a, U5b, U5b1c1, U5b2a1a, V, W, X
- Salzmünde group (5,400 to 5,000 ybp : central-east Germany): H (x2), H3 (x2), H5, HV, HV0, J, J1c (x2), J2b1a, K1, K1a, K1a4a1a2, N1a1a1a3 (x2), T2b (x2), U3a, U3a1, U5b, V, X2b1’2’3’4’5’6
- Outliers from Gotland, Sweden (5,300 to 4,700 years ago): H7d, HV0a, J1c5 (2x), J1c8a, K1a2b (2x), K2b1a, T2b8
- Middle Neolithic Portugal (c. 5,800 to 5,400 years ago): H, H1, H10e, HV0, J (x2), K1a2a1, T2b, U4, U5, U5b
- Globular Amphora Culture (c. 4,850 to 4,450 years ago: Poland): K, K2a ref
Sauveterrian, Tardenoisian, and contemporary cultures (10,500 to 6,500 years ago; West & Southwest Europe)
- Chandler et al. (2005) recovered 8 mtDNA sequences from several Mesolithic sites from the Sado estuary in central Portugal, and identified 4 individuals belonging haplogroup H (including one H1b and one possible H7), 2 to haplogroup U (U4 and U5b1c2) and 2 to haplogroup N (probably N1b and N5). Their age range from 9,500 to 7,500 years ago.
- Delsate et al. (2009) analyzed the mtDNA of the Reuland-Loschbour man (8,000 years ago) from Luxembourg and assigned him to haplogroup U5a.
- In 1996, Bryan Sykes of Oxford University first sequenced the mitochondrial DNA of the 9,000-year-old Cheddar Man from Gough’s Cave in Cheddar (Somerset), England, and assigned him to haplogroup U5.
- Hervella et al. (2012) tested one mtDNA samples from Aizpea (Navarre) dated from 6,600 years ago, and found it to belong to haplogroup U5b1. ref
Maglemosian-Kongemose, Kunda-Narva, Neman-Zedmar, and related cultures (11,500 to 5,000 years ago; North & Northeast Europe)
- Bramanti et al. (2009) tested Mesolithic remains from several locations across Europe, and found one haplogroup U5a (9,800 years ago) at the Chekalino site in the Volga-Ural region of Russia, one U5a1 (10,000 to 8,000 years ago) at the Lebyazhinka site in the Middle Volga region of Russia, one U5b2 (9,200 years ago) at the Falkensteiner Höhle cave in Baden-Württemberg, Germany, one U5a2a and one U5b2 (both 8,700 years ago) at the Hohlenstein-Stadel cave, in Baden-Württemberg, Germany, one U4 (8,850 years ago) at Bad Dürrenberg in Saxony-Anhalt, Germany, one U4 and one U5b2 (both 8,350 years ago) from the Kunda Culture in Lithuania, two U5b2 (both 6,450 years ago) from the Narva Culture in Lithuania, two U5b1 (6,000 to 5,000 years ago) from the Zedmar Culture in Poland and one U5a (4,250 years ago) from the Drestwo site in northeast Poland.
- Balanovsky et al. (2012) tested 19 ancient mtDNA sequences from Gotland, Sweden. They identified 8 individuals belonging haplogroup U4, 6 to haplogroup U5 (including three U5a), one to haplogroup V, one to haplogroup K, one to haplogroup T. No haplogroup could be attributed for the last 2 samples based on the HVR test alone (16311C). ref
Pitted Ware Culture (5,200 to 4,000 years ago; Scandinavia)
- Malmström et al. (2009) tested 19 ancient mtDNA sequences from Gotland, Sweden. They identified 8 individuals belonging haplogroup U4, 6 to haplogroup U5 (including three U5a), one to haplogroup V, one to haplogroup K, one to haplogroup T. No haplogroup could be attributed for the last 2 samples based on the HVR test alone (16311C). Rivollat et al. (2020) tested one additional Pitted Ware sample who belonged to Y-haplogroup I2a2a1b and mt-haplogroup JT. ref
Starčevo–Kőrös–Criş Culture and Linear Pottery Culture (c. 8,000 to 6,500 years ago; Central & Southeast Europe)
- Haak et al. (2005) and Haak et al. (2010) sequenced the mitochondrial DNA from several LBK sites in Germany and one in Austria dating from 5500 BCE to 4900 BCE. Out of the 38 mtDNA lineages recovered there were six haplogroup N (one N1a, one N1a1a, two N1a1a1, two N1a1a2, and one N1a1b), two U (U3 and U5a1a), seven K, four J, ten T (including three T2), three HV, eight H, two V, and two W. The Y-chromosomal DNA of three samples was also successfully retrieved and assigned to haplogroup F* (2 samples) and G2a3.
- Bramanti et al. (2008) tested the mtDNA from the LBK site of Vedrovice (5300 BCE) in the Czech Republic. Two samples were found to belong to haplogroup K, one to J1c, two to T2 and the last one to H.
- Guba et al. (2011) analyzed the mtDNA of 11 Neolithic skeletons from Hungary. Among the five specimens from the Kőrös culture (5500 BCE), two carried the mutations of haplogroup N9a and one of C5. Another one had a series of mutations not seen in any haplogroup to this day (16235G, 16261T, 16291T, 16293G, 16304C). The last one didn’t have any mutation from the CRS in the HVS-I region and is, therefore, undetermined. Out of the six specimens from the LBK-related Alföld Culture (5250-5000 BCE) three belonged to haplogroup N (N1a, N1a1b, N9a), and one to haplogroup D1 or G1a1. The two others were undetermined (CRS and 16324C mutation reported as M/R24). ref
Cucuteni-Trypillian Culture (c. 7,500 to 4,750 years ago; Romania, Moldova, Ukraine)
- Nikitin et al. (2010) studied the remains of the Eneolithic site of Verteba Cave (3600-2500 BCE) in Western Ukraine. They retrieved the mtDNA of seven individuals, which were assigned to haplogroup pre-HV, HV or V (2 samples), H (2 samples), J and T4. ref
Cardium Pottery Culture (c. 8,400 to 4,700 years ago; Mediterranean Europe)
- Chandler et al. (2005) sequenced the mtDNA of four Neolithic skeletons from the Impressed Ware Culture of Portugal (5500-4750 BCE), and found two members of haplogroup U (U and U5), one of H and one of V.
- Lacan et al. (2011) tested 29 skeletons from a 5,000-year-old site in Treilles, Languedoc, France. Twenty paternal lineages (Y-DNA) were identified as G2a, while the two others belonged to haplogroup I2a. The maternal lineages (mtDNA) comprised six haplogroup U (including four U5 and one U5b1c), two K1a, six J1, two T2b, two HV0, six H (three H1 and three H3), one V, and four X2. The two I2a men belonged to mtDNA haplogroup H1 and H3.
- Lacan et al. (2011 bis) tested 7 skeletons from a 7,000-year-old Neolithic site from the Avellaner Cave in Cogolls, Catalonia, Spain. Six paternal lineages (Y-DNA) were identified including five G2a and one E1b1b1a1b (E-V13). There were three mtDNA haplogroup K1a, two T2b, one H3, and one U5.
- The team of Fernández et al. (2006) and Gamba et al. (2008) analyzed the mitochondrial HVR-I in 37 bone and teeth samples from 17 archaeological sites located around Castellón de la Plana, Valencia, Spain. Most of the results were inconclusive though. Out of the 12 mtDNA sequences from the Chalcolithic period that were retrieved, four were reported as haplogroup L3, four as H (including three CRS, which could be non-results), two to R0, HV or H, one to V, and one to D.
- Gamba et al. (2011) identified the mtDNA of 10 Early Neolithic (5000-5500 BCE) samples from the sites of Can Sadurni and Chaves and three Late Early Neolithic (4250-3700 BCE) from Sant Pau del Camp, all around Barcelona, Spain. The coding region was also tested to confirm the haplogroups. The results included haplogroups N* (4 samples), H (4 samples including one H20), U5 (1 sample), K (3 samples), and X1 (1 sample). ref
Atlantic Megalithic Culture (c. 7,000 to 4,000 years ago; Western Europe)
- Deguilloux et al. (2010) examined skeletons from the Péré tumulus, a megalithic long mound (4200 BCE) in Brittany, and retrieved the mtDNA of three individuals. They belonged to haplogroup N1a, U5b and X2.
- Sampietro et al. (2007) analyzed the HVRI mitochondrial DNA sequences of 11 Neolithic remains from the Cami de Can Grau site (3500 BCE) in Granollers, Catalonia, Spain. Four skeletons belonged to haplogroup H (including three CRS, which could be non-results), two to J, two to T2, one to U4, one to I1, and one to W1.
- In a study focusing mostly on the site of Tell Halula in Syria, Fernández et al. (2008) also tested two skeletons from the Nerja caves near Málaga, Andalusia, Spain. The first individual (3875 BCE) carried the mutations 16126C 16264T 16270T 16278T 16293G 16311C, and the second 16129A, 16264T, 16270T, 16278T, 16293G, 16311C. Both sequences could correspond either to haplogroup H11a (typical of Central Europe) or, more probably L1b1 (found in the Canaries and Northwest Africa).
- In one pioneering ancient DNA study N. Izagirre and C. de la Rua (1999) of the University of the Basque Country, analysed the mtDNA variations in 121 dental samples from four Basque prehistoric sites. Among them, 61 samples from the late Neolithic site of San Juan Ante Portam Latinam (3300-3042 BCE) in Araba were found to belong to haplogroups H (23 samples), J (10 samples), U (11 samples), K (14 samples) and T or X (3 samples). The site of Pico Ramos (2790-2100 BCE) in Bizkaia yielded 24 results including haplogroups H (9 samples), J (4 samples), U (3 samples), K (4 samples), and T or X (4 samples). The site of Longar (2580-2450 BCE) in Nafarroa had 27 individuals H (11 samples), U (4 samples), K (6 samples), T or X (4 samples) and two other unidentified haplogroups. Finally, the site of Tres Montes in Navarra (2130 BCE) possessed 3 samples that appeared to belong to haplogroup L2 and two others that were undetermined (16224C and 16126C+16311C). The authors noted the conspicuous absence of haplogroup V, now present at a relatively high frequency among the Basques (6.5%).
- Fernández et al. (2005) tested the mtDNA of remains from the Abauntz site (2240 BCE) in Navarra. All three samples retrieved were inconclusive regarding the mitochondrial haplogroup. One sample was CRS (no mutation found). Another had 16126C+16311C, which would be R0a, HV0a, or a subclade of H, among many other possibilities. The last one (16256T) could be H1x, H14, or even U5a.
- Hervella et al. (2012) tested 48 mtDNA samples from northern Spain dating from 6,185 to 5,160 years ago. Among the 41 Early Neolithic samples from Los Cascajos and Paternanbidea (Navarre), they observed 2 members of haplogroup H3 (or possibly H1a1), 8 H, HV or R0 (CRS), 4 other H, 1 HV, 1 U5, 8 U, 4 K, 2 J, 1 T2, 1 I and 1 X. Among the remaining seven Middle Neolithic samples, one from Marizulo (Gipuzkoa) belong to U5, while the six from Fuente Hoz (Alava) belonged to H (2, including one rCRS) and U (4 samples, including one U5a). ref
Funnelbeaker Culture (c. 6,000 to 4,700 years ago; Northern Europe)
- Malmström et al. (2009) tested three mtDNA sequences from a megalithic site (3500-2500 BCE) in Gökhem, Sweden. They identified haplogroups H, J, and T.
- Bramanti et al. (2009) tested seven skeletal materials from Ostorf (3200-3000 BCE) in Mecklenburg-Vorpommern, Germany, and identified the haplogroups as U5 (3 samples including one U5a), K and T2e (2 samples) and J. ref
Bell Beaker phenomenon (c. 4,400 to 3,800 years ago; Western Europe)
- Melchior et al. (2010) managed to retrieve two mtDNA sequences from the Damsbo site (2200 BCE) in Denmark, which belonged to haplogroups U4 and U5a2a.
- Lee et al. (2012) retrieved the DNA of eight skeletons (6 in individual graves and 2 in double graves) in a cemetery located at Kromsdorf, Thuringia, Germany. The samples were dated between 2600 and 2500 BCE. Six mtDNA sequences were obtained and attributed to haplogroups U2e, U5a1, T1a, K1, I1, and W5a. The Y-DNA of two skeletons was also tested and assigned to haplogroup R1b (including one positive for M269, but negative for the Germanic S21/U106). ref
Corded Ware (Battle-axe) Culture (c. 4,900 to 4,300 years ago; North and Northeast Europe)
- Haak et al. (2008) extracted the mitochondrial DNA of 7 skeletal materials from a grave in Eulau (2,600 BCE). The remains belonged to mitochondrial haplogroups H, U5b, K1a2, and K1b (2 samples), I and X2. The Y-chromosomal DNA of two of the men was successfully extracted and found to belong to haplogroup R1a1. ref
Western European Bronze Age (c. 4,500 to 2,800 years ago; Western Europe)
- Kerri Brown of the University of Manchester retrieved the DNA from various bones of two Bronze Age skeletons from the Hebrides in Scotland, known as the Cladh Hallan mummies (1500 BCE). She explained that the mummies were made of body parts from several different people, arranged to look like one person. The mitochondrial DNA retrieved was reported on TV as H, U, or U5, and probably T1.
- N. Izagirre and C. de la Rúa (1999) tested 6 Bronze-age mtDNA from the Basque site of Urratxa (circa 1450 BCE) in Bizkaia, Spain. They found two specimens belonging to haplogroups H, two to J, and two to U.
- Simon et al. (2011) retrieved the DNA from eight individuals in collective burials (circa 1200 BCE) at the Montanissell Cave in the Catalan pre-Pyrenees, Spain. The coding region was tested for two CRS results that turned out to be haplogroups U and J. They managed to assign four individuals to haplogroup J (three or all four probably J1c), two to haplogroup U (including one U5a1c), one to haplogroup K, and the last one to haplogroup HV0 or V.
- Hervella et al. (2012) tested 2 mtDNA samples from Urtiaga (Gipuzkoa), which were found to belong to haplogroups H, HV, or R0 (rCRS) and U5a. ref
Urnfield Culture (c. 3,300 to 2,750 years ago; Central Europe)
- Felix Schilz (2006), of the University of Göttingen, extracted the DNA of 34 skeletons from the Lichtenstein Cave (1,000 BCE) in the Harz mountains of central Germany. Using short tamdem repeats (STR) variances, 11 male individuals were assigned to Y-DNA haplogroup I2b2, two to haplogroup R1a, and the last one to R1b. On the maternal side, 16 samples belonged to mitochondrial haplogroups H, 5 to haplogroup J (including one J1b1), 5 to haplogroup T2, 8 to haplogroup U (including five U5b and one possible U2). ref
Nordic Bronze Age (c. 3,700 to 2,500 years ago; Scandinavia)
- Melchior et al. (2010) recovered the mtDNA of a skeleton from Bredtoftegård site (circa 1400 BCE) in Denmark. He was identified as a member of haplogroups U4. ref
Balkanic Bronze Age (c. 4,500 to 3,200 years ago; Southeast Europe)
- Cardos et al. (2004) analyzed the mitochondrial DNA from 5 Bronze-age individuals from Southeast Romania. Only a few HVR-1 mutations were successfully identified, which is not enough to determine unambiguously their haplogroups. The first specimen had the mutation 16129C. Assuming that no other HVR-1 mutation is missing due to incomplete sequencing or damaged DNA, this would fit haplogroup H17. The second individual had the mutations 16186T, 16189C and 16299T, which could be either H1a or H39 with two private mutations. The next person carried mutation 16129A and 16223T, which could also be haplogroup H1j or H17 with a private mutation, or even HV4a2 with 16221 missing. The fourth sample had only 16145A, which defines H22 with 16227 (it could therefore be “pre-H22”). The last sample had the common 16311C mutation, which if it is the only difference from the CRS would correspond to H2b, but is also found in subclades of H3, H6, H11, H13, and all subclades from HV6 to HV11. In conclusion, if all the HVR region was thoroughly tested and didn’t miss any mutation (a big ‘if’), then all the samples probably belonged to various subclades of H. However missing several mutations, they could belong to almost anything. ref
Mycenaean Greece (c. 3,900 to 3,100 years ago; Greece)
- Bouwman et al. (2008) tested the mitochondrial DNA of 22 skeletons from Grave Circle B at Mycenae (1,500 BCE). They only managed to identify 4 sequences, which belonged to haplogroups H (CRS), U5a1a, K (2 samples, probably brother and sister). ref
Nuragic civilisation(c. 3,800 to 1,850 years ago; Sardinia)
- The team of Caramelli et al. (2007) and Ghirotto et al. (2010) tested 23 HVR-1 mtDNA sequences from Bronze Age Sardinia (ranging from 1430 to 930 BCE) to compare them to modern Sardinian sequences. They found 11 ancient samples belonging probably to haplogroup H (including 6 CRS, which could be non-result), one HV0 (reported as V), two U2 (or possibly H1a3), one J, three samples with the mutation 16129C that can correspond to H1j, H17 or possibly even U, and two samples with the mutation 16223T that do not permit to assign a haplogroup unambiguously. ref
Srubna culture, Novocherkassk culture, and Cimmerian civilization (c. 3,800 to 2,450 years ago; Pontic Steppe)
- Nikitin et al. (2011) tested the mtDNA of remains from the Verteba Cave in central Ukraine. The first sample (circa 700 BCE) was attributed to haplogroup U5a1a. The second one (circa 750 BCE) matched the mutations for haplogroup H5a. ref
Iron Age (c. 3200 to 1500 years ago)
Dacian and Thracian civilizations (c. 3,000 to 1,900 years ago; Southeast Europe)
- Cardos et al. (2004) tested 4 bone remains from the Babadag Culture (1000-600 BCE) in Romania. Like for the Bronze-age samples, none of the results were conclusive though. One sample didn’t have any HVR-1 mutation differing from the CRS. One sample had the mutations 16192T and 16284C. The two other samples both had 16262C, a mutation that isn’t part of any known European or Middle Eastern haplogroup. ref
Iberian civilization (3,100 to 2,200 years ago, in eastern Spain)
- Sampietro et al. (2005) tested the mtDNA of 17 skeletal remains from Iberian sites dating from 700 to 100 BCE scattered around Girona in Catalonia, Spain. To ascertain the haplogroups, the team analysed a few key mutations in the coding region in addition to the hypervariable section, and were able to determine that 9 individuals belonged to haplogroup H (including two H1), one R0, two J*, one T, three U (one U4, one U5 and one U5a), and one K.
- The team of Fernández et al. (2006) and Gamba et al. (2008) analyzed the mitochondrial HVR-I in 37 bone and teeth samples from 17 archaeological sites located around Castellón de la Plana, Valencia, Spain. Among the 8 mtDNA sequences from the Iron Age (700-500 BCE) that were retrieved, three individuals were reported as members of haplogroup H (all three CRS, so inconclusive), one V, one T1, one K, and two W. ref
Scandinavian Iron Age (500 BCE to 1,000 CE)
- Melchior et al. (2008a) analyzed the mtDNA sequences of 7 skeletons from Bøgebjerggård (c. 2,000 years ago), 6 from Simonsborg (c. 1900 years ago) and 11 from Skovgaarde (c. 1,750 years ago), all in Denmark. They found 10 members of haplogroup H (including one H1), one V, one R0a, two J, one T2b, four U (one U2e, one U3a, one U5b, one U5b1), two K and three I.
- Melchior et al. (2008b) sequenced 10 Viking skeletons from Galgedil in Denmark (c. 1000 years ago). They found 5 members of haplogroup H (including one H5a1g), one I, one K, one T2, four U5a1a, and one X2c.
- Naumann et al. (2013) tested the mtDNA of 9 Norwegian Norsemen from Flakstad in Norway (c. 1000 years ago) and reported five members of haplogroup H (including one H1a), two J (including one J1b1), one U4, and one U5a.
- Krzewińska et al. (2014) sequenced 43 Viking Age Norwegian skeletons from Hedmark, Nordland, Nord-Trøndelag, Oppland, Sør-Trøndelag, Telemark and Troms (c. 1000 years ago). They found 16 members of haplogroup H (including one H1b, H2a1 and one H6), two HV0, two HV (HV4a and HV9a), two I (including one I1), six J (including three J1 and one J1b1), four K (including one K1a11), one T2b, one U3a, one U4, seven U5 (including U5a and one U5b1b1), and one V. ref
Lombards (500-800 CE)
- Alt et al. (2014) analyzed the DNA of 45 skeletons from a 6th-century Lombard cemetery at Szólád in Hungary, and managed to recover 24 mtDNA sequences. They found 8 members of haplogroup H (including one H1c1 and one H2a2b), two HV, two I3, four J (including one J1b1a and one J2b), two K, one N1a, one N1b2, two T2b (including one T2e), one U* and three U4.
- Vai et al. (2015) sequenced 102 Lombard skeletons from 5 burial sites in Northwestern Italy and successfully obtained 28 mitochondrial sequences. They found 14 members of haplogroup H (including one H2a2b1, three H2b, one H6a1b1, and two H24), one I2a, five J (including one J2a2c), three T (including two T2e), one U2e1, one U4, and three U5a (two U5a1b1e and one U5a2a). ref
“Researchers present the integrative bioarchaeological study on the Pre-Pottery Neolithic B (PPNB) in Southeastern Anatolia by combining isotopic data (87Sr/86Sr, δ18O, and δ13Ccarb), new radiocarbon dates, and genome-wide data recovered from human skeletal remains from the site of Nevalı Çori. We also report human genome-wide data from post-Neolithic Nevalı Çori and the late PPNB site of Ba’ja in the Southern Levant. Our combined isotope and ancient DNA data fill a research gap between prehistoric Anatolian and Levantine populations. Our results indicate a decline in human mobility after the first phase of the PPNB in the Southeastern Anatolia accompanied by increasing reliance on domesticated resources and evidence of consanguinity in the PPNB Levant.” ref
“Growing reliance on animal and plant domestication in the Near East and beyond during the Pre-Pottery Neolithic B (PPNB) (the ninth to eighth millennium BCE) has often been associated with a “revolutionary” social transformation from mobility toward more sedentary lifestyles. We are able to yield nuanced insights into the process of the Neolithization in the Near East based on a bioarchaeological approach integrating isotopic and archaeogenetic analyses on the bone remains recovered from Nevalı Çori, a site occupied from the early PPNB in Turkey where some of the earliest evidence of animal and plant domestication emerged, and from Ba’ja, a typical late PPNB site in Jordan. In addition, we present the archaeological sequence of Nevalı Çori together with newly generated radiocarbon dates.” ref
“The study’s results are based on strontium (87Sr/86Sr), carbon, and oxygen (δ18O and δ13Ccarb) isotopic analyses conducted on 28 human and 29 animal individuals from the site of Nevalı Çori. 87Sr/86Sr results indicate mobility and connection with the contemporaneous surrounding sites during the earlier PPNB prior to an apparent decline in this mobility at a time of growing reliance on domesticates. Genome-wide data from six human individuals from Nevalı Çori and Ba’ja demonstrate a diverse gene pool at Nevalı Çori that supports connectedness within the Fertile Crescent during the earlier phases of Neolithization and evidence of consanguineous union in the PPNB Ba’ja and the Iron Age Nevalı Çori.” ref
“Overall, these variegated connections in material culture and genetics point to different spheres of interactions between Anatolia, the Levant, and the Upper Mesopotamia. The genetic variability among populations from Mesopotamia suggests that the range from Southern to Eastern Anatolia and Northern Iraq contained different niches of blending ancestries that could have variably contributed to other populations in the Levant and the rest of Anatolia. Another genetic evidence, the ROH estimated on NEV009 and Çayönü individuals indicate that these individuals belonged to a population of a larger effective population size compared with the coeval individuals from the Southern Levant, Western Iran, and Southcentral Anatolia. This corroborates different demographics in Upper Mesopotamia, which could have promoted long-range mobility resulting in many of the observed genetic signals in ancestry. Additional genetic data from the neighboring areas would be critical for investigating the overall dynamic transition that SE Anatolia experienced during the PPNB.” ref
Disconnection and Collapse of the TSP Horizon.
Genetic Study Detects Unexpected Origin of World’s First Farmers
“Did the hunter-gatherers of Anatolia have an epiphany and beget the earliest farmers, or were they somebody else entirely? About 10,000 years ago, the story of humanity reached a turning point, transiting from a life of hunting and gathering to subsistence agriculture and animal husbandry (with some hunting and gathering). This transition – the “Neolithic revolution” – appeared at dierent times in dierent parts of the world (and never even reached Australia, where farming really only began in the 19th century). Agriculture and animal husbandry apparently developed independently in dierent areas during the Holocene.” ref
“But leaving China out of it, one of the very first places agriculture and the practice of breeding captive herbivores emerged was the region of Anatolia-Mesopotamia. The Neolithic revolution changed the trajectory of the human story. The question is: who exactly were the early farmers of Anatolia? Were they locals living there from time immemorial who developed a new lifestyle? Invaders? Something else? All of the above?” ref
“Now, analysis of ancient DNA samples from all over the Near East sheds light on that conundrum, report Drs. Iosif Lazaridis, Songül Alpaslan-Roodenberg, Ron Pinhasi and David Reich of Harvard University, and a giant international team, in Science. The early farmers were not pure locals. There were two pulses of migration into Anatolia during the early Neolithic. The first had already occurred by the Pre-Pottery Neolithic period, about 11,000 to 9,000 years ago. It originated in northern Mesopotamia. “The Mesopotamian admixture is present in all Pre-Pottery Neolithic Anatolians we sampled, but not in an Epipaleolithic individual from Pinarbasi in Anatolia from about 15,500 years ago (which is the one hunter-gatherer datapoint from Anatolia),” Reich claries.” ref
“The second migration into early Neolithic Anatolia distinguishes all the Anatolian farmers from the Pottery Neolithic period that began about 9,000 ago from those that preceded them. The source was the Levant, Lazaridis explains. In other words, the incomers didn’t supplant or extinguish the locals; they admixed. Thus, the early farmers of Anatolia have three distinct deep hunter-gatherer ancestries: Anatolian, Mesopotamian and Levantine, Lazaridis sums up. Actually, all the sampled Neolithic populations throughout Middle East are a mixture of these three deep sources, Lazaridis says. “It’s probably more complicated,” Reich qualies. But for the nonce, we can describe the early farmers of Anatolia as mixes, in dierent proportions, of these three source populations.” ref
Wishing well?
“One challenge with studying our origins is the paucity of material. One might think Africa and Eurasia are littered with human remains from our entire evolutionary history, but they aren’t. Even after deep burial emerged, as opposed to shallow burials vulnerable to hyenas, no burial at all or cremation, precious little is preserved over time. Altogether, the paper is based on genomic analysis of just 100 ancient individuals: 42 individuals with new data, and 48 previously published. From Israel, for example, the team analyzed DNA from two groups: six pre-Neolithic Natuans from about 13,000 years ago, and two Pre-Pottery Neolithic people from about 9,000 years ago. The Cyprus data, which is the first ever reported from this island, was based on the fragmentary remains of three folk who, about 10,000 years ago, were apparently tossed down a well.” ref
“Tsk tsk. From Anatolia, the study analyzes 49 individuals – including three newly reported individuals from the Pre-Pottery Neolithic site of Boncuklu Tarla in Mardin, southeast Turkey. Along with two individuals from Nemrik in Iraq dating to a similar time, these represent the first ancient DNA data ever reported from Mesopotamia. The study also reports two individuals from Armenia who lived around 8,000 years ago – the first Neolithic data from the Armenian plateau – and several more from the previously unsampled northern Zagros mountain range from Iraq’s Shanidar and Bestansur caves.” ref
“It would be nice to have more samples but such is life – the sheer number of reports based on analyses of ancient genomes tends to obscure the extraordinary diculty of the technique. But the samples suced to deduce that before the Neolithic, there were distinct hunter-gatherer populations in Anatolia, Mesopotamia (the Caucasus) and the Levant. Come the Neolithic, the people had become mixes, with varying proportions of the three sources in dierent places, Reich and Lazaridis explain. Put otherwise, none of these populations – not in Anatolia or in the Levant or southern Mesopotamia – descended from just one of these three sources, Lazaridis stresses. But the Neolithic people of the Levant have more Natuan than other sources, for example.” ref
“All this is from the team’s analysis of ancient DNA. There is also some concrete evidence of contact between these peoples, such as an obsidian blade from Anatolia found in Motza (by Jerusalem) in a 9,000-year-old village, and another found in prehistoric Saudi Arabia. Of course, these blades don’t necessarily attest to direct relations; they could have changed hands over generations as they slowly wound their way from Turkey to Israel and Arabia. That Neolithic village in Motza is also where one of the two Levantine samples in this study was from; the other was from Kfar Horesh. Both were from the Pre-Pottery period, and genetically, they seem almost identical to the parallel population in ‘Ain Ghazal in Amman, Jordan, Reich adds.” ref
“How large were these two pulses of migration? We have no idea, but they weren’t a trickle. The mixes are showing up as 30 percent, 40 percent, 50 percent – which suggests much movement, the researchers explain. But they can’t say whether these were “sudden” processes over just a few generations, or a protracted process of exchanges over millennia, Reich says. There just isn’t enough data at this point. “With richer sampling, we’d begin to learn this. In Britain we have tons of sampling and can pinpoint population change to a few centuries, but we don’t have that here,” he adds.” ref
Anatolians in Israel
“One wonders at the scantiness of information from Israel, which sometimes seems to have more archaeologists than people. “Israel is the place in the Near East with the richest data and the archaeology is incredible,” Reich responds. But so far, most attempts to retrieve DNA from skeletal remains in Israel have been frustrating – as said, the technique is a monster, conditions in Israel aren’t necessarily conducive to adequate preservation of DNA in ancient bones, and thus managing to extract DNA from Natuan remains at Rakefet Cave was practically a miracle. However, the technology is improving, Reich says. Meanwhile, Lazaridis points out that they don’t have data on the Pottery Neolithic period from the Levant (8,000 to 9,000 years ago), so can’t be condent the migration was bidirectional. Come the Chalcolithic (the Copper Age), there is a “beautiful set of 22 samples” from northern Israel’s Peki’in cave, with extra Anatolian ancestry, Reich says. One site with evidence of Anatolian ancestry in northern Israel 6,000 years ago does not a mass movement make, but it’s intriguing. Then there’s a gap in the genetic information on the peopling of the Levant from about 6,000 to 4,000 years ago, Lazaridis notes. Not surprisingly, they have no data from any period from Syria or Lebanon.” ref
“What the new information can’t do is shed light on the rise and fall of the people who brought us Göbekli Tepe, Karahan Tepe, and altogether 16 sites (discovered so far) in prehistoric Turkey. The thinking now is their construction began about 12,000 years ago, by pre-Neolithic hunter-gatherers, denitely not early farmers, says Prof. Necmi Karul of Istanbul University. These hunter-gatherers were not living in small nomadic groups: they had villages and built great monuments that some call “the earliest temples in the world,” and that Karul prefers to call “gathering places.” After about 1,500 years, that culture seems to have disappeared. But there are no pre-Neolithic samples from that area and only one from Anatolia (that person from Pinarbasi who lived about 15,000 years ago), Lazaridis says. At this point, they can’t answer the question of continuity between pre-Neolithic and Neolithic populations in that part of the world.” ref
“Also, as Reich points out, Göbekli wasn’t a burial site. “These burials are scarce, especially in the hunter-gatherer period. It’s the rare region where there is a cemetery or cave context. We have every hope of getting such data in future, but for now we don’t have it,” he says. What we do have is a better picture of the early farmers in the Near East: they were a mix, not pure descendants of local hunter-gatherers, insofar as that could be tested. Where the Levant component originated is not clear; the southern Levant – say, Jordan or Israel, or perhaps Syria – from where there is no data, but where there had been a rich Pottery-period Neolithic culture. The team adds some caveats about using the word “migration.” “When talking about “migration pulses,” it likely wasn’t intentional; likely no master plan of conquest was involved; the
dimensions are unclear. As they say, there aren’t many samples, but a pattern is a pattern – and Homo genus seems to have had wanderlust since we had feet.” ref
Körtiktepe
“Körtiktepe, also known as Körtik Tepe, is the oldest known Neolithic site in Turkey. Together with Tell Mureybet and Tell Abu Hureyra in northern Syria, Körtiktepe is one of the only three securely dated Younger Dryas sedentary sites in Upper Mesopotamia. The habitation of the site began in the first half of the 11th millennium BCE, approximately 10700 BCE (12,700 years ago), and persisted with consistent density until approximately 10400 BCE. Strontium and oxygen isotope analyses of human tooth enamel indicate that the inhabitants of the Younger Dryas occupations at Körtiktepe were born and grew up in or near the site. Although a potential minor flooding event transpired during the transition from the Younger Dryas to the Early Holocene, the site endured without evident abandonment, at least not for a prolonged interval. Occupation continued and thrived during the Early Holocene. The architectural tradition of constructing round plans established around 10400 BCE and continued without any fundamental alterations until the eventual desertion of the site. The site reached its peak in terms of occupation density around 9300 BCE. Subsequently, it experienced an unexplained abandonment, possibly attributed to natural disturbances such as flooding induced by the Holocene climate changes.” ref
“Abstract: Recent archaeological discoveries, refinements in genetic analyses, and the archaeobotanical data require a reconsideration of the nature of the emergence of sedentary farming communities in Southeastern Turkey. In the rescue excavations prompted by the Batman and Ilısu dam projects, several Early Holocene sites were discovered. For the first time, new data from Körtik Tepe now provide detailed evidence of a local Epipalaeolithic origin for these permanent settlements. In this article, we summarize the results and analyses of the 2010-2012 excavations as well as palaeopedological and archaeobotanical data of the Younger Dryas layers at Körtik Tepe. Human isotope studies and the archaeological data suggest a permanent occupation of the site and might point to a local primordial adoption of a sedentary lifestyle in this region as early as the 11th millennium BCE.” ref
Çemka Höyük
“Çemka Höyük, in the Upper Tigris Basin, has the potential to provide important information on PPNA and Late Epipaleolithic Period cultures in this region. The excavations to be carried out in the following years shed light not only on the Neolithization process of the region in question but also on broader interactions between regions. Çemka Höyük is a settlement that provides important information on many points such as the transition from the Late Epipaleolithic to the PPNA (especially hunter-gatherer semi-nomadic life) and the development of the Zarzian culture in the north, especially the Late Epipaleolithic Period.” ref
“In recent studies carried out at the Çemka Höyük settlement on the western flank of the Tigris River in Upper Mesopotamia (Mardin province, Turkey), new information has been obtained on the Late Epipaleolithic and Early Neolithic periods, defined as the Proto-Neolithic. Despite the short-term nature of the excavations, the settlement provides new data about these eras in the region; in particular, the settlement is significant in terms of the Late Epipaleolithic–Neolithic transition as well as of architectural finds belonging to both periods. The rise of permanent settlements and domestic architecture is a focus of examination.
“A “shaman” burial from the PPNA settlement of Çemka Höyük, Upper Tigris Basin, Turkiye. Knowledge of the burial customs of the Pre-Pottery Neolithic A (PPNA) in the Near East is increasing. Particularly, lately a large number of burials and skeletal remains have been unearthed in the Upper Tigris Basin, thanks to a number of new excavation projects in recent years. The newly revealed findings indicate that PPNA burial customs varied considerably in the region from site to site. However, the 10th millennium BCE burial ÇH 2019/05 at Çemka Höyük shows as well that there are also different burial practices with in settlements. ÇH 2019/05 belongs to a female individual, accompanied by animal skeletal elements, who appears to may have been a shaman or at least had been buried by someone practicing ways associated with what we understand nowadays as animism or shamanism. Hence, the burial may represent one of the earliest known examples of its kind in an Anatolian Neolithic context.” ref
“Human remains from Near Eastern Neolithic sites show regional and chronological differences in sepulchral organization (Bocquentin et al., 2016). However, few Near Eastern sites have provided information on burial customs for the PPNA. For example, if one does not take into account the human remains unearthed at sites set up in the Upper Tigris Valley, excavated in previous years, there are few human remains in the Near East as a whole for the PPNA. Human remains from Körtik Tepe, Hasankeyf Höyük, Gre Fılla, Demirköy, Çayönü, Gusir Höyük, Boncuklu Tarla, and Çemka Höyük provide important information for the burial customs of the Upper Tigris region, and also those of the Near East.” ref
“At these sites, primary burials are in the majority, and skulls taken from Demirköy and Boncuklu Tarla are rarely found. Skeletons with traces of paint are characteristic at Körtik Tepe and Hasankeyf Höyük. At the same time, the absence of human remains at the Hallan Çemi site shows that there are differences within the same region. In contrast, human remains are rare in northern Syria. There are a few human skeletons in Tell Qaramel, Jerf el Ahmar, Dja’de, Tell Qaramel, and Mureybet. In eastern Jezirah, the site of Nemrik 9 has yielded a number of human remains related to primary and multiple burials and human remains related to secondary burials at Qermez Dere. However, the practice of skull removal (isolated skulls and skull-less skeletons) and isolated human bones are found in a variety of architectural contexts; in or outside houses, in community buildings, etc, in the Middle Euphrates Valley, at Göbekli Tepe; in Northern Syria at Dja’de, Tell Abr 3 and Jerf el-Ahmar; Mureybet, Tell Qaramel; in Iraqi Jezirah at Nemrik 9 (Kozlowski, 1990) and Qermez Dere, as well as in the southern Levant.” ref
“Indeed, funerary gestures are varied during the 10th millennium throughout the northern Near East. Above all, human remains are found in great quantities at some sites, such as Körtik Tepe, and at others, they are rarely found or not found at all. We also notice different modes of burial such as primary, secondary, skull-less skeletons or removed skulls. In this regard, a cattle skull and a number of cattle bones, together with the remains of a small ruminant, a partridge bone, a canid bone, and a marten bone, associated with a PPNA burial in Çemka Höyük — dating to the end of the 10th millennium BCE — provide new insights into animal-human relationships at the dawn of the Neolithic in the Upper Tigris Valley. The finds and animal remain associated with the human burial context raise questions about the relationship between PPNA burial customs and the use of especially cattle skulls, and adds new data to the discussion on human-cattle relationship in the early Neolithic of Northern Mesopotamia.” ref
Woman buried 12,000 years ago in Turkey may have been a shaman
“A small team of archaeologists in the Republic of Türkiye has identified a woman who was buried approximately 12,000 years ago in a neolithic settlement now called Çemka Höyük as a possible shaman. At Çemka Höyük, in the southeastern part of Türkiye, discovered the remains of a woman dating to approximately 12,000 years ago—a time when the people living there were still hunter–gatherers, but who had also begun establishing small settlements. The researchers believe she was between the ages of 25 and 30 when she died. The woman had been buried under the floor of a building constructed of mud. Prior research has shown that burying people under their homes was a common practice of the time. But unlike other contemporaneous remains, she was found buried with a large variety of animals. At the time of her death, domestication of animals had not yet begun; thus, they were all wild. Perhaps even more surprising is that the woman’s body was covered by a large block of limestone.” ref
“After careful study of the animals buried with the woman showed that they had been placed carefully—the skull of an aurochs had been placed just above her body, for example, with its jaw separated and placed at her feet. Other bones included those of a partridge’s wings, a marten’s legs and random sheep or goat bones scattered throughout the burial plot. The researchers suggest burying animals with the woman, a practice not seen with others, hints at the possibility that the woman held a special place in society, perhaps working as a shaman—a person who would have had a special connection with spirits, especially those with evil intent. Shamans in other societies have often been perceived as having a close connection with animals and the spirits that may inhabit them.” ref
Two New Dogs, and Other Natufian Dogs, from the Southern Levant
“Abstract: Two new fairly complete remains of dogs were uncovered from a burial in Hayonim Terrace (northern Israel), dated to the first half of the 11th millenium years ago (Late Natufian). This burial contained the remains of three humans associated with an elaborate construction. A detailed analysis of these dogs, and a comparison with all known Natufian remains, suggested that genuine dogs were already living around and within human habitations during this period. While most studies on early dogs were carried out on post-Natufian material, a period during which intentional selection could have already been widely experienced, we show that the evolvement of Natufian dogs were the product of unconscious selection of commensal wolves quasi-isolated under the special anthopogenic habitats created within and around Natufian sites, and at least ritually assimilated to human society. There is no evidence that Neolithic dogs are direct descendants of Natufian ancestors. Their multiregional origination is a widely accepted phenomenon. We suggest that Neolithic dogs were either domesticated anew, or were introduced from elsewhere to the southern Levant. Although the number of specimens of Natufian dogs is still small, evidently the emergence of some patterns clearly indicate that all them were already markedly different from the recent southern Levantine wolves and, in spite of their widely ranged morphology, constituted a unique group. It is shown that the shortening of the muzzle mainly affected the anterior part of the snout, while the posterior region remained practically unchanged. In this respect they seem to display a typical case of paedomorphosis. The simultaneous diminution of the carnassials, and other teeth, alongside with the snout, was marked enough that no crowding of the teeth took place by any of the Natufian dogs. The disproportional reduction of the snout versus teeth that caused the “crowding” phenomenon is only known from later periods.” ref

ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref
This art above explains my thinking from my life of investigation
I am an anarchist (Social anarchism, Left-wing anarchism, or Socialist anarchism) trying to explain prehistory as I see it after studying it on my own starting 2006. Anarchists are for truth and believe in teaching the plain truth; misinformation is against this, and we would and should fight misinformation and disinformation.
I see anarchism as a social justice issue not limited to some political issue or monetary persuasion. People own themselves, have self/human rights, and deserve freedoms. All humanity is owed respect for its dignity; we are all born equal in dignity and human rights, and no plot of dirt we currently reside on changes this.
I fully enjoy the value (axiology) of archaeology (empirical evidence from fact or artifacts at a site) is knowledge (epistemology) of the past, adding to our anthropology (evidence from cultures both the present and past) intellectual (rational) assumptions of the likely reality of actual events from time past.
I am an Axiological Atheist, Philosopher & Autodidact Pre-Historical Writer/Researcher, Anti-theist, Anti-religionist, Anarcho Humanist, LGBTQI, Race, & Class equality. I am not an academic, I am a revolutionary sharing education and reason to inspire more deep thinking. I do value and appreciate Academics, Archaeologists, Anthropologists, and Historians as they provide us with great knowledge, informing us about our shared humanity.
I am a servant leader, as I serve the people, not myself, not my ego, and not some desire for money, but rather a caring teacher’s heart to help all I can with all I am. From such thoughtfulness may we all see the need for humanism and secularism, respecting all as helpful servant leaders assisting others as often as we can to navigate truth and the beauty of reality.
‘Reality’ ie. real/external world things, facts/evidence such as that confirmed by science, or events taken as a whole documented understanding of what occurred/is likely to have occurred; the accurate state of affairs. “Reason” is not from a mind devoid of “unreason” but rather demonstrates the potential ability to overcome bad thinking. An honest mind, enjoys just correction. Nothing is a justified true belief without valid or reliable reason and evidence; just as everything believed must be open to question, leaving nothing above challenge.
I don’t believe in gods or ghosts, and nor souls either. I don’t believe in heavens or hells, nor any supernatural anything. I don’t believe in Aliens, Bigfoot, nor Atlantis. I strive to follow reason and be a rationalist. Reason is my only master and may we all master reason. Thinking can be random, but reason is organized and sound in its Thinking. Right thinking is reason, right reason is logic, and right logic can be used in math and other scientific methods. I don’t see religious terms Animism, Totemism, Shamanism, or Paganism as primitive but original or core elements that are different parts of world views and their supernatural/non-natural beliefs or thinking.
I am inspired by philosophy, enlightened by archaeology, and grounded by science that religion claims, on the whole, along with their magical gods, are but dogmatic propaganda, myths, and lies. To me, religions can be summed up as conspiracy theories about reality, a reality mind you is only natural and devoid of magic anything. And to me, when people talk as if Atlantis is anything real, I stop taking them seriously. Like asking about the reality of Superman or Batman just because they seem to involve metropolitan cities in their stores. Or if Mother Goose actually lived in a shoe? You got to be kidding.
We are made great in our many acts of kindness, because we rise by helping each other.
NE = Proto-North Eurasian/Ancient North Eurasian/Mal’ta–Buret’ culture/Mal’ta Boy “MA-1” 24,000 years old burial
A = Proto-Afroasiatic/Afroasiatic
S = Samara culture
ST = Proto-Sino-Tibetan/Sino-Tibetan
T = Proto-Transeurasian/Altaic
C = Proto-Northwest Caucasus language/Northwest Caucasian/Languages of the Caucasus
I = Proto-Indo-European/Indo-European
IB = Iberomaurusian Culture/Capsian culture
Natufian culture (15,000–11,500 years ago, Syria, Lebanon, Jordan, the Sinai Peninsula, and the Negev desert)
Nganasan people/Nganasan language
Na-Dene languages/Dené–Yeniseian, Dené–Caucasian
Proto-Semitic/Semitic languages
24,000 years ago, Proto-North Eurasian Language (Ancient North Eurasian) migrations?
My thoughts:
Proto-North Eurasian Language (Ancient North Eurasian) With related Y-DNA R1a, R1b, R2a, and Q Haplogroups.
R1b 22,0000-15,000 years ago in the Middle east creates Proto-Afroasiatic languages moving into Africa around 15,000-10,000 years ago connecting with the Iberomaurusian Culture/Taforalt near the coasts of Morocco, Algeria, and Tunisia.
R2a 10,000 years ago in Iran brings/creates Proto-Indo-European language and also a possibility is R1a in Russia around 9,000 years ago may have had a version of Proto-Indo-European language.
Around 14,000-10,000 years ago??? Proto-North Eurasian Language goes to the Yellow River basin (eventually relating with the Yangshao culture) in China creates Proto-Sino-Tibetan language.
Proto-Sino-Tibetan language then moves to the West Liao River valley (eventually relating with the Hongshan culture) in China creating Proto-Transeurasian (Altaic) language around 9,000 years ago.
N Haplogroups 9,000 years ago with Proto-Transeurasian language possibly moves north to Lake Baikal. Then after living with Proto-North Eurasian Language 24,000-9,000 years ago?/Pre-Proto-Yeniseian language 9,000-7,000 years ago Q Haplogroups (eventually relating with the Ket language and the Ket people) until around 5,500 years ago, then N Haplogroups move north to the Taymyr Peninsula in North Siberia (Nganasan homeland) brings/creates the Proto-Uralic language.
Q Haplogroups with Proto-Yeniseian language /Proto-Na-Dene language likely emerge 8,000/7,000 years ago or so and migrates to the Middle East (either following R2a to Iraq or R1a to Russia (Samara culture) then south to Iraq creates the Sumerian language. It may have also created the Proto-Caucasian languages along the way. And Q Haplogroups with Proto-Yeniseian language to a migration to North America that relates to Na-Dené (and maybe including Haida) languages, of which the first branch was Proto-Tlingit language 5,000 years ago, in the Pacific Northwest.
Sino-Tibetan language then moves more east in China to the Hemudu culture pre-Austronesian culture, next moved to Taiwan creating the Proto-Austronesian language around 6,000-5,500 years ago.
R1b comes to Russia from the Middle East around 7,500 years ago, bringing a version of Proto-Indo-European languages to the (Samara culture), then Q Y-DNA with Proto-Yeniseian language moves south from the (Samara culture) and may have been the language that created the Proto-Caucasian language. And R1b from the (Samara culture) becomes the 4,200 years or so R1b associated with the Basques and Basque language it was taken with R1b, but language similarities with the Proto-Caucasian language implies language ties to Proto-Yeniseian language.
Genetic evidence for two founding populations of the Americas
“Abstract: Genetic studies have been consistent with a single common origin of Native American groups from Central and South America. However, some morphological studies have suggested a more complex picture, whereby the northeast Asian affinities of present-day Native Americans contrast with a distinctive morphology seen in some of the earliest American skeletons, which share traits with present-day Australasians (indigenous groups in Australia, Melanesia, and island southeast Asia). Here, we analyze genome-wide data to show that some Amazonian Native Americans descend partly from a Native American founding population that carried ancestry more closely related to indigenous Australians, New Guineans, and Andaman Islanders than to any present-day Eurasians or Native Americans. This signature is not present to the same extent or at all in present-day Northern and Central Americans or a ~12,600-year-old Clovis genome, suggesting a more diverse set of founding populations of the Americas than previously accepted.” ref
Karitiana
“The Karitiana or Caritiana are an indigenous people of Brazil, whose reservation is located in the western Amazon. They count 320 members, and the leader of their tribal association is Renato Caritiana. They subsist by farming, fishing and hunting, and have almost no contact with the outside world. Their tongue, the Karitiâna language, is an Arikém language of Brazil. A 2015 genetic study reached a surprising conclusion about the origins of the Karitiana people. While the Karitiana people are closely related to other Native Americans, they share closer relations to Southeast Asians & Polynesians compared with other Native Americans which are closest to Siberians and Northeast Asians.” ref
“A study by Iosif Lazaridis (2014) found Karitiana to carry Mal’ta MA1 (41%) admixture while the other geneflow in Karitiana appears to have an Eastern Eurasian origin. A study by Kanazawa-Kiriyama et al. (2017) detected gene flow from Karitiana to Mal’ta MA1 (21%) which is in the reverse direction of what was reported in previous studies such as Raghavan et al. 2014 who used a much larger sequence data. The authors speculate that the inverse flow could be due to Ancient Beringian migration in a westward migration into Eurasia. Studies of population genetics often use the Karitiana as a reference population for Native Americans, using DNA samples made available through the Human Genome Diversity Project and other sources.” ref
“DNA from Karitiana individuals was collected in 1987 by Francis Black and in 2007 it was reported that this sampling was undertaken unbeknownst to FUNAI, the Brazilian agency that regulates contact between the indigenous tribes and the outside world, and that the samples were being distributed for a fee with no benefit to the Karitiana, giving rise to claims of biopiracy. The same newspaper report claimed that further samples were taken in 1996 by Dr. Hilton Pereira da Silva, a doctor on a documentary film crew, on the promise of medicinal supplies that were never fulfilled. A response from Dr. Silva suggests that the news story was faulty and the medicinal samples he took were never used for any commercial purpose.” ref
“Current research trends in the Paleolithic studies have the major objective of providing answers to questions concerning the time of origin, characteristic features, and developments of Stone Age archaeological cultures. Apparently, priority is given to the studies of the morphology and typology of stone tools as well as to the technology of their production. The apparent predominance of this trend is stipulated by the fact that lithic artifacts represent the principal and, in most cases, sole items of anthropogenic origin at the majority of Paleolithic archaeological sites. Other information, including that of the original conditions of artifact deposition within soft sediments, is often missing due to natural, primarily geological processes. The overwhelming majority of Paleolithic sites older than 20,000 years are located on piedmont shelves, alluvial fans, and terrace-like escarpments of mountain slopes. Because they are located in these geological contexts, a considerable number of archaeological sites have been impacted by the destructive influence of denudation processes.” ref
“In such situations, detailed studies of the preserved structural elements of occupation sites have gained special importance. The whole diversity of structural elements can be subdivided into simple (hearths, spots of burned soil, household pits, hoards, dwelling zone pavements, refuse pits, stone and bone knapping zones, ash concentration zones, etc.) and complex (dwelling places possessing a combination of simple elements) features. Studies of these elements allow researchers to reconstruct the original structural patterns at Paleolithic sites. This research can also provide information on the behavioral strategies of early populations, not only with respect to economic activities, but also in the spiritual sphere. In 1991, the Podzvonkaya Paleolithic site was discovered in the eastern part of the Kyakhta Region of the Buryatia Republic. Podzvonkaya revealed well-preserved structural elements of the spatial organization of an ancient occupation site.” ref
“At first, the site consisted of a rather small area that yielded archaeological materials attributed to the Upper Paleolithic. The site is located in an amphitheater formed by the hills on the left bank of the narrow valley of the Tamir River in the western ranges of the Tamir Mountain Range. Archaeological excavations carried out from 1991 – 2000 revealed that the site is wider than the initially established area of 100 x 50 m. At present, the entire territory of the south-facing piedmont shelves, ranging from the floodplain to the eastern slopes of the amphitheater, has been identified as the early Upper Paleolithic occupation zone. The total area of the site is divided into several portions by the topography of the amphitheater. In total, four loci with stratified archaeological and paleontological materials have been investigated. These loci have been designated as the East, West, Southeast, and Lower Complexes. The East and Southeast Complexes are of major interest in the studies of spatial patterning of an early occupation site. Focusing on hearths and spots of burned soil from culture-bearing horizon 3 of the East Complex. The author provides an interpretation of specific features of hearths and associated osteological materials in their archaeological contexts.” ref
“The data discussed in the present paper suggest two major conclusions. 1. The Podzvonkaya Paleolithic site was repeatedly occupied by humans, possibly representing a seasonal camp. The noted structural elements of the site allow us to infer that representatives of a single population group inhabited this site during several seasons. The Podzvonkaya site was occupied for a rather long period in the early Upper Paleolithic, during which the earlier structural elements were covered by refuse left behind by succeeding generations of inhabitants. 2. The repeated recovery of finds demonstrating features, either identical or similar in particular aspects, from hearths and fireplaces at Podzvonkaya, together with specific features of deposition of the artifacts, suggest their premeditated placement into the hearths. This fact is suggestive of intentional actions performed by Paleolithic humans with respect to hearths and fireplaces when they were no longer in use.” ref
“Apparently, these actions were ritual and were aimed at places of burning fire rather than at the fire itself, because some bones and skeletal fragments were placed in hearths after the fire was extinguished. The abovementioned materials recovered from the hearths and fireplaces from the East Complex at Podzvonkaya are not unique evidence of early ritual behavior in Trans-Baikal. For instance, the Paleolithic site at Varvarina Gora yielded a cult burial of animal bones: a skull of a carnivore and horse bones were recovered from a small pit with a stone pavement at the bottom. These facts allow us to infer that Paleolithic humans felt the necessity to comprehend the phenomena of the surrounding world and tried to establish a relevant behavior strategy in the form of spiritual interrelationships with this world. Let us propose a chronological estimate of the period of hearth use during which the rituals of hearth cult were performed. The Podzvonkaya lithic industry is based on the sub-prismatic and prismatic stone reduction strategy with a considerable proportion of Levallois flaking. Available archaeological materials illustrate a trend of development of the Upper Paleolithic tool types (lateral racloirs, knives, points on blades, burins, and adzes). The set of distinctive Middle Paleolithic tool types includes dejete side- scrapers, asymmetrical Mousterian points, etc.” ref
“The noted combination of the major tool types suggests an early Upper Paleolithic attribution of the site. Available radiocarbon dates do not contradict such age estimations. A date of 38,900 ± 3300 years ago (AA-26741) has been generated on a bone sample from the lithological level containing Hearth 1 of the East Complex. This date is well correlated to a series of dates from other Complexes at the site and dates the whole site back to about 40,000 years ago. In this case, the site is one of the oldest representations of the hearth cult and ritual actions aimed at hearths that occurred in the southern West Trans-Biakal in the early Upper Paleolithic. Archaeological investigations carried out at other portions of the Podzvonkaya site have revealed hearth cult representations similar to those noted at the East Complex. Other excavation areas have shown a poor state of preservation of layers containing archaeological materials. For instance, the Southeast Complex revealed hearth remains destroyed by solifluction processes. The hearth possessed a pavement of water-worn stone slabs and yielded a fragment of a horse mandible and bones of an extremity of an artiodactyl. The extremity bones were grouped together, and some of them preserved anatomical order. Similar finds were recovered from the Lower Complex. Thus, the evidence suggests a single cultural tradition.” ref
Afanasievo culture
“The Afanasievo culture, or Afanasevo culture (Afanasevan culture) (Russian: Афанасьевская культура Afanas’yevskaya kul’tura), is an early archaeological culture of south Siberia, occupying the Minusinsk Basin and the Altai Mountains during the eneolithic era, c. 3300 to 2500 BCE. It is named after a nearby mountain, Gora Afanasieva (Russian: Гора Афанасьева, lit. ‘Afanasiev’s mountain’) in what is now Bogradsky District, Khakassia, Russia, first excavated by archaeologist Sergei Teploukhov in 1920-1929. Afanasievo burials have been found as far as Shatar Chuluu in central Mongolia, confirming a further expansion about 1,500 km beyond the Altai Mountains. The Afanasievo culture is now considered as an integral part of the Prehistory of Western and Central Mongolia.” ref
“According to David W. Anthony the Afanasevan population was descended from people who migrated c. 3700–3300 BCE across the Eurasian Steppe from the pre-Yamnaya Repin culture of the Don–Volga region. It is considered as “intrusive from the west”, in respect to previous local Siberian cultures. According to Anthony, “The Afanasievo culture migration to the Altai was carried out by people with a Repin-type material culture, probably from the middle Volga-Ural region.” A 2021 study by F. Zhang and others found that early Tarim mummies from the late 3rd and early 2nd millennia BCE were unrelated to the Afanasevians, and came from a genetically isolated population derived from Ancient North Eurasians, that had borrowed agricultural and pastoral practices from neighboring peoples.” ref
“Because of its geographical location and dating, Anthony and earlier scholars such as Leo Klejn, J. P. Mallory and Victor H. Mair have linked the Afanasevans to the Proto-Tocharian language. Afanasievan ancestry persisted in Dzungaria at least until the second half of the 1st millennium BCE. The Shirenzigou culture (410–190 BCE), just northeast of the Tarim Basin, also appears to have been derived from the Afanasievans, which, in addition to linguistics, further reinforces an Afanasievo hypothesis for the Tocharians. The first Afanasievo archaeological site was found near the mountain of Gora Afanasieva (Minusinsk Basin). It was excavated in 1920-1929 by Russian archaeologist Sergei Teploukhov, and the newly discovered culture was named after the mountain.” ref
“The original Afanasievo site was on the first floodplain terrace of the Yenisei river near Gora Afanasieva, 1 km to the southeast from the village of Bateni-Yarki, and is now submerged in the flood zone of the Krasnoyarsk Reservoir since 1960-67. Many other Afanasievo sites were found in the Ukok Plateau, and as far south as the area around Ürümqi (Tuqiu), near the Tarim Basin, and the area of Dzungaria. The area from the Minusinsk Basin to Dzungaria is the main area of Afanasievo occupation, but recently, Afanasievo burials were found as far east as Altan Sandal and Shatar Chuluu in central Mongolia, confirming an eastward expansion about 1,500 km beyond the Altai Mountains and beyond the previously known area of occupation.” ref
“Conventional archaeological understanding tended to date the Afanasievo culture at around 2500–2000 BCE. However radiocarbon gave dates as early as 3705 BCE on wooden tools and 2874 BCE on human remains. The earliest of these dates have now been rejected, giving a date of around 3300 BCE for the start of the culture, and 2500 BCE for its termination. Mass graves were not usual for this culture. Afanasievo cemeteries include both single and small collective burials with the deceased usually flexed on their back in a pit. The burial pits are arranged in rectangular, sometimes circular, enclosures marked by stone walls. It has been argued that the burials represent family burial plots with four or five enclosures constituting the local social group.” ref
“The Afanasievo economy included cattle, sheep, and goat. Horse remains, either wild or domestic, have also been found. The Afanasievo people became the first food-producers in the area. Tools were manufactured from stone (axes, arrowheads), bone (fish-hooks, points) and antler. Among the antler pieces are objects that have been identified as possible cheek-pieces for horses. Artistic representations of wheeled vehicles found in the area has been attributed to the Afanasievo culture. Ornaments of copper, silver and gold have also been found. The Afanasievans are now considered as the earliest herders of East Asia, who were instrumental in the establishment of the long tradition of pastorialism in Mongolia.” ref
“Their rise also corresponds with the appareance of domesticated sheep, goats, and cattle, which marks the earliest spread of Near Eastern domesticated animals and pastoralism to Inner Asia. They also introduced the initial practice of copper and bronze metallurgy. Afanasievo burials include metal artifacts in copper, bronze (awls, knives), gold and silver, as well the remains of disassembled carts. The Afanasievos may have used cattle-drawn wagons, as did Yamnaya communities. Petroglyphs of animals are associated to the area and period of the Afanasievo culture and share similarities with petroglyphs found in western and central Asia. Animal hunting, 3000 BCE, Arkhangai Province, Mongolia. National Museum of the Altai Republic.” ref
“The analysis of the full genome of Afanasievo individuals has shown that they were genetically very close to the Yamnaya population of the Pontic–Caspian steppe. The Afanasievo and Yamnaya populations were much more similar to each other than to groups geographically located between the two (which, unlike Afanasievo samples, carried a large amount of ancestry from eastern Siberian hunter-gatherers). This indicates that the Afanasievo culture was brought to the Altai region via migration from the western Eurasian steppe, which occurred with little admixture from local populations. From the Altai mountains, steppe-derived Afanasievo ancestry spread to the east into Mongolia and to the south into Xinjiang.” ref
“The Yamnaya-related lineages and ancestry in Afanasievo disappeared in the course of the Bronze Age in the Altai region and Mongolia, being replaced by the migrating populations from the Sintashta culture arriving from the west. In Dzungaria, Afanasievo-related ancestry persisted at least into the late first millennium BCE. The Afanasievo people, accompanied by their pastoralist technologies, are one of the major foreign contributors to the genetic profile of the modern northwestern Chinese. The genetic closeness of the Yamnaya and Afanasievo populations is also mirrored in the uniparental haplogroups, especially in the predominance of the Y-chromosome haplogroup R1b. A 2018 study analyzed the maternal haplogroups of 7 Afanasievo specimens. 71% belonged to West Eurasian maternal haplogroups U, H, and R, while 29% belonged to the East Eurasian maternal haplogroup C.” ref
“Afanasievo burials are recorded as far as central Mongolia, at the sites of Altan Sandal and Shatar Chuluu. To their east, in the modern area of eastern Mongolia and beyond, resided Neolithic cultures of Prehistoric Mongolia, probably derived from the Ancient Northeast Asians, who were the predecessors of the Slab Grave culture of eastern Mongolia. At Afanasevo Gora, two strains of Yersinia pestis have been extracted from human teeth. One is dated 2909–2679 BCE; the other, 2887–2677 BCE. Both are from the same (mass) grave of seven people, and are presumed near-contemporary. This strain’s genes express flagellin, which triggers the human immune response; so it was not a bubonic plague.” ref
“Because of its numerous traits attributed to the early Indo-Europeans, like metal-use, horses and wheeled vehicles, and cultural relations with Kurgan steppe cultures, the Afanasevans are believed to have been Indo-European-speaking. They were genetically similar to the Yamnaya populations of Western Steppe Herders. Genetic studies have demonstrated a discontinuity between Afanasievo and the succeeding Siberian-originating Okunevo culture, as well as genetic differences between Afanasievo and the Tarim mummies. A genomic study published in 2021 found that the population of earliest Tarim Basin cultures (the Tarim mummies, dated to c. 2000 BCE) had high levels of Ancient North Eurasian ancestry and no connection with Afanasievo populations.” ref
“Numerous scholars have suggested that the Afanasievo culture may be responsible for the introduction of metallurgy to China. In particular, contacts between the Afanasievo culture and the Majiayao culture and the Qijia culture are considered for the transmission of bronze technology. The Afanasievo culture may also display cultural borrowings from the earlier Banpo culture (c. 4000 BCE), particularly in the area of painted pottery, suggesting influence from the Far East, specifically from Neolithic China, on the Afanasievo culture and other cultural complexes in the Middle Yenisei region. According to Allentoft and coauthors (2015): “Afanasievo culture persisted in central Asia and, perhaps, Mongolia and China until they themselves were replaced by fierce warriors in chariots called the Sintashta (also known as the Andronovo culture).” ref
“In the Altai Mountains and to the southeast, Afanasievans seem to have coexisted with the early period of the Chemurchek culture for some time, as some of their burials are contemporary and some of the artifacts of the burials coincide. To the north, the Afanasievo culture was succeeded by the Okunev culture, which is considered as an extension of the Paleosiberian local non-Indo-European forest culture into the region. The Okunev culture nevertheless displays influences from the earlier Afanasievo culture. The region was subsequently occupied by the Andronovo, Karasuk, Tagar and Tashtyk cultures, respectively. Allentoft et al. (2015) confirmed that the Afanasevo culture was replaced by the second wave of Indo-European migrations from the Andronovo culture during late Bronze Age and early Iron Age. The Andronovo population was found to be genetically related, but clearly distinct from the Afanasievo population.” ref
“Several scholars propose the Afanasievo culture as the ancestors of the Tocharians, who lived on the northern edge of the Tarim Basin (in present-day Xinjiang, China) in the first millennium CE. The Tocharian languages are believed to have become extinct during the 9th century CE. The Indo-European speaking Tocharian peoples of the Tarim city-states then intermixed with the Uyghurs, whose Old Uyghur language spread through the region. Genetic studies on Iron Age individuals of the Shirenzigou site dated to circa 200 BCE have shown a fairly balanced admixture between the West Eurasian and East Eurasian genetic pools.” ref
“The West Eurasian component was Yamnaya-related, while the East Eurasian component was Northeast Asian-related. The Yamnaya component suggest a strong probability that the Shirenzigou populations were derived from the Afanasievo culture to the north, and spoke an Indo-European language. This reinforces an Afanasievo hypothesis for the Tocharians, often called the “Steppe hypotheses”, rather than an hypotheses favouring BMAC and Andronovo Culture origins, the “Bactrian Oasis hypotheses”. Allentoft et al. (2015) sampled four females from the Afanasievo culture, two individuals carried mtDNA haplogroup J2a2a, one carried T2c1a2, and one carried U5a1a1. Narasimhan et al. (2019) analyzed the remains of 24 individuals ascribed to the Afanasievo culture. Of the 14 samples of Y-DNA extracted, 10 belonged to R1b1a1a2a2, 1 to R1b1a1a2a, and 3 belonged to Q1a2. The mtDNA samples belonged to subclades of U (particularly of U5), along with T, J, H, and K.” ref
Okunev culture
“Okunev culture (ru: Окуневская культура, romanized: Okunevskaya kul’tura, lit. ‘Okunev culture’), also known as Okunevo culture, was a south Siberian archaeological culture of pastoralists from the early Bronze Age dated from the end of the 3rd millennium BCE to the early 2nd millennium BCE in the Minusinsk Basin on the middle and upper Yenisei. It was formed from the local Neolithic Siberian forest cultures, who also showed evidence of admixture from Western Steppe Herders and pre-existing Ancient North Eurasians.” ref
“Okunev culture was discovered by Sergei Teploukhov in 1928. It was named after the nearby Okunev settlement in the south of modern day Khakassia. Initially, the burials from Okunev were attributed by Teploukhov to the Andronovo culture. Then, on the basis of vessel finds, Teploukhov considered the population to be a transitional variant between the Afanasievo and Andronovo cultures. In 1947, M. N. Komarova identified an “early Okunev” stage of the Andronovo culture, which is associated with the earliest stage of the Andronovo culture.” ref
“In 1955-1957 A.N. Lipsky found Okunev stone slabs with images as part of stone boxes used for burials. Lipsky, who was an ethnographer, not an archaeologist, assumed that the Okunev sites were pre-Afanasiev and attributed them to the Paleolithic era, since he considered the Okunev people to be the ancestors of the American Paleo-Indians, based on parallels in art and anthropology. In the early 1960s G. A. Maksimenkov identified an Okunev culture based on the excavations of the Chernovaya VIII burial ground, whose burials had not been disturbed by later invasions and did not contain Afanasevo ceramics.” ref
“The early Uibat stage, later Chernov stage, and the final Razliv stage of Okunev culture need to be differentiated. Typical sites include Tas-Khaaz, Beltyry, Uibat III, Uibat V (in the Uybat river basin), Chernovaya VIII, Chernovaya XI, Razliv X, and Strelka. The typological horizon between the development of the Afanas’ev and Okunev steppe cultures in the Minusinsk Basin and the development of the later Andronovo type is very thin. Finds from the Okunev culture include works of art, including stone statues with human faces (Tas Khyz, as well as Ulug Khurtuyakh tas) and images of birds and beasts hammered out on stone slabs or engraved on bone plaques. There are no significant indications of property and social stratification.” ref
“The basis of the population’s economic activity was stock-raising and animal husbandry (cattle, sheep, and goats), supplemented by hunting and fishing. Stone hoes, grain graters, and pestles, and a reaping sickle with a copper blade and horn handle all testify to agriculture. Though the ceramic styles of the Okunev are more comparable to later Incised Coarse Ware (ICW), formally and ambiguously Andronovo ceramics. But as the researchers note, the uniqueness of each of them is an important feature of the Okunev culture. Finds from the Okunev culture include lavishly decorated jug-like and conical vessels. Okunev ceramics are typically flat-bottomed, with notable continuous ornamentation of the body, the bottom, edge of the rim and its inner side. Most often these are jar vessels, but there are also incense burners with an internal partition.” ref
“Okunevtsy had developed metallurgy based on the ores of the Sayano-Altai mining and metallurgy areas. Okunevtsy and the neighboring Samus culture produced the first bronze in north-eastern Central Asia. Finds include copper and tin and rarely arsenical bronze articles. Simple copper objects were superseded by tin alloys. Bronzes were common in this culture. Tools included embedded-handled knives, leaf-shaped knives, awls, fishhooks, and temporal rings. Along with forging, casting was also used, which indicates a rather high level of metalworking. Ornaments of this culture consist mainly of ring-shaped ornaments with circular cross-sections and flat joints at both ends.” ref
“Short swords are relatively advanced with clear boundaries between the handles and the blades. A bronze spear was found at the late Okunevo cultural site, the socket of which was forged with two loose ends. The first of this kind appeared in the Asian steppe region. Besides copper and bronze weapons, the Okunev culture also had charriots as attested by their petroglyphs. The Okunevo culture is represented mostly by mounds burial structures, which were composed of small, rectangular surface enclosures made of stone slabs or sandstone tiles placed vertically in the ground. Within these enclosures were graves that were also lined with stone slabs. 62 Okunevo kurgans consisting of more than 500 burials and 60 single burials have been studied.” ref
“The cemeteries of the Okunev culture are located, as a rule, not far from the Afanasiev ones and number from two to ten burial mounds. Sometimes burial complexes measure 40 × 40 meters. The number of graves inside the fence varies – from one to ten and even twenty. In addition to single burials, there are paired and collective burials. In almost every burial ground there are burials of a man with two women. The buried were laid, as in Afanasiev’s time, on their backs with legs strongly bent at the knees and arms extended along the body. Radiocarbon AMS dating of 50 Okunevo samples are within 2600 –1800 BCE. According to these studies the Uybat period is dated as 2600 – 2300 BCE, Chernovaya as 2200 – 1900 BCE, and Razliv later than 1800 BCE.” ref
“The settlements of the Okunev culture were located in the Minusinsk basin, on the middle and upper Yenisei. Petroglyphs of the Okunev period are located in the narrow mountain valleys of Khyzyl Khaya and Khurtuy Khola, on the banks of now-dry streams in the modern Kazanovka Museum-Reserve. They are also represented among the Shalabolino Petroglyphs on the right bank of the Tuba River, a right tributary of the Yenisei, against the village of Tes to the southeast of the village of Ilyinka, between the logs and the Shush River to the southwest of the village of Shalabolino, Kuraginsky District, Krasnoyarsk Krai. In the Idrinsky district, east of the village of Bolshoi Telek.” ref
“In the Krasnoturansky District on the left bank of the Bir River under Mount Kozlikha, on the banks of the Syda River. In the Kuraginsky district in the village of Novopokrovka. In the Minusinsky District, on the banks of the Tuba River, near the village of Kavkazsky, nearby the zaimka of Maidashi. On the shore of the lake near the village of the same name Maly Kyzykul, during excavations in the Okunev layer, archaeologists in 1973 discovered the remains of a burnt log structure and fragments of ceramic dishes. Five burials in slab boxes were excavated 1 km south of Minusinsk on the northern outskirts of a pine forest.” ref
“Okunev culture shares some elements of its material culture, including pottery. with a number of local contemporaneous cultures from adjacent areas such as the Samus’, Elunino, Karacol, and Krotovo cultures of western Siberia and Altai, the Kanay type burials of eastern Kazakhstan, and the Okunevo-like culture of Tuva. Nevertheless, there is currently no sound evidence of their common origin. The connections between the Afanasiev and Okunev cultures are rather difficult to trace. The period of their interaction lasted only about a hundred years, but in some territories coexistence is noted. Archaeologists have identified many complexes containing signs of both Okunev and Afanasevo origins. However, almost no genetic traces of Afanasevtsy have been found in the Okunev genotype, meaning Afanasiev population was displaced by the alien Okunevtsy.” ref
“The similarity between some of the objects from the Okunev burial grounds and objects in the vicinity of the middle Ob River and the Lake Baikal region indicates that the bearers of the Okunev culture came to southern Siberia from the northern taiga regions. While the preceding Afanasevo culture is considered Indo-European, the Okunev culture is generally regarded as an extension of the local non-Indo-European forest culture into the region. The Okunev people closely interacted with successor cultures of the Andronovo circle. The settlements of this culture have been little studied. Mountain Fortress Sve mountain settlements with fortifications (about 45 were found on the territory of Khakassia) are mainly considered cult complexes. The fortress of Chebaki is one of the first archeologically studied Sve.” ref
“The Okunev people used two- and four-wheeled carts. In the rock art of the Minusinsk Basin, images of early (end of the 3rd millennium BCE) two-wheeled carts with a composite drawbar of two poles converging at an angle, which simultaneously form the body frame, are common. The design of the wagons and the profile manner of depiction indicate a connection not with Eastern Europe, but with the western regions of Central Asia and, indirectly, with Asia Minor. The anthropological type of the population was of mixed Caucasoid-Mongoloid origin, with a predominance of Mongoloid. As A. V. Gromov notes, their morphological heterogeneity was striking – there are both purely Mongoloid skulls and typically Caucasoid skulls that do not reveal any traces of Mongoloid admixture. In his opinion, the appearance of the Okunev people was formed as a result of the mixing of the local Neolithic population with an influx of Afanasyevtsy from the territory of Central Asia and Kazakhstan.” ref
“According to A. G. Kozintsev, the appearance of the Okunev people varies depending on the region. The Okunev people of the Minusinsk Basin were descendants of the local Neolithic population, which was distinguished by its significant originality against the background of the races of the first order. The Okunev people of Tuva show stronger influence from the Pits culture and early Catacomb culture of Ukraine. He argues that the main ancestry of the Okunev people can be traced back to the local Ancient North Eurasians (ANE) and that the anthropologic type of the Okunev people can be described as “Americanoid“, noting the specific overlaps in characteristics with the Indigenous peoples of the Americas. According to A. V. Polyakov, the culture was formed from the local Neolithic Paleo-Siberian forest cultures and later received some admixture from the Caspian Sea by a group of mostly male pastoralists of the Yamnaya culture.” ref
“While some authors have suggested that the Okunevo may have descended from more northern tribes that replaced Afanasievo cultures in this region, others believe the Okunevo culture was the result of contact between local Neolithic hunter-gatherers with western pastoralists. Maksimentkov suggested that Okunevo culture was developed by the local Neolithic tribes of the Krasnoyarsk – Kansk forest-steppe who lived to the north of the Minusinsk Basin. The second theory that is supported at the present time by most researchers suggests that Okunevo culture resulted from the interaction of local Neolithic hunter-gatherers with Western Steppe Herders.” ref
“Autosomal DNA analysis found that the Okunevo people formed predominantly from a lineage originating from the admixture of Ancient Northeast Asians (ANA) with Ancient North Eurasians (ANE), with around 10-20% genetic admixture from Western Steppe Herders, as represented by the Yamnaya or Afanasievo cultures. The Western Steppe Herder ancestry is absent from the X chromosome of Okunevo spcecimens, suggesting it was inherited from mostly male ancestors. The date of admixture is estimated to have been around 7,000 years ago. According to recent studies, modern Native American Indians are genetically close to representatives of the Okunev culture, which confirms previous craniometric studies. Their shared affinities probably come from the presence of Ancient North Eurasian and Ancient East Asian ancestries in both populations dating back to the formation of Ancient Paleo-Siberians.” ref
“The Okunevo population showed also genetic affinities with the Botai culture, some of the Tarim mummies, and Altai hunter-gatherers. The results of the analysis of the origin of the ancient steppe populations of nomads of the Eurasian steppe (from the Urals to Altai), including representatives of the Bronze Age Okunev culture from the Sayan-Altai, showed that the samples contained components that were most pronounced in Ancient North Eurasian, Eastern hunter-gatherers, Caucasian hunter-gatherers from Georgia and also occur from the component that is most pronounced among the Nganasans (Samoyedic people) and is widely distributed among various modern people from Siberia and Central Asia.” ref
“Hollard et al. (2018) reported the paternal haplogroups of 6 Okunevo specimens. 50% of the Okunevo males belonged to the East Eurasian haplogroup NO(xO). The other 50% belonged to East Eurasia originated West Eurasian haplogroups: including 33% assigned to Q1b, and 16% with R1b1a2-M269. According to Holllard (2018), 58% of Okunevo specimens carried the East Eurasian haplogroups A, C or D, while 41% carried the West Eurasian haplogroups T, U, H or J. The mitochondrial haplogroup A-a1b3* was identified in the RISE674 sample (4300–3850 years ago, Okunevo_EMBA). In representatives of the Okunev culture from the burial ground of Syda V (Minusinsk Basin), a variety of mitochondrial DNA variants was determined. The Okunevs belonged to the West Eurasian (U, H, J, and T) and East Eurasian (A, C, and D) subbranches of haplogroups.” ref
“Representative art: small amulets, stone steles up to 4 m tall, and petroglyphs. The Okunev people left behind monuments of art. Characteristic rock inscriptions and stone statues have become famous since the travels of D. G. Messerschmidt in 1722-1723 and subsequent academic expeditions. Steles with drawings from burial vaults are unique. The stone slabs are dominated by realistic images of animals and masks in headdresses, which apparently had a cult character. Rock art monuments are being studied and new ones are being discovered that were not studied by previous researchers. Menhirs are common in the territory of modern Khakassia and the southern part of the Krasnoyarsk Krai. More than 300 of them have been explored on the territory of the Minusinsk Basin. Only 10 sites are known on the right bank of the Yenisei.” ref
The impressive stone steles were originally erected at gravesites and were subsequently reused more than a millennium later in the Scythian-era kurgans of Tagar Culture. Okunev stone stela collections are displayed in the Khakassia National Museum in Abakan, Martyanov Museum in Minusinsk, Historical and architectural open-air museum of Novosibirsk and ceramics collections are displayed in the Hermitage Museum in Saint Petersburg. The vivid character of the art of the Okunev culture is created by monumental stone sculptures and steles carved with anthropomorphic images. The stone statues are usually tall, up to six meters in height, carved of sandstone or granite into a saber shape. The front is its narrow edge. More than 300 of them have been studied in the Minusinsk Basin, cur only ten are known on the right bank of the Yenisei river. Many of them are now in museums.” ref
“A fantastic mask looks at the viewer from it: three eyes, nostrils, a huge mouth, horns, long ears, and all kinds of processes. The image moves from the front face to the wide side, and sometimes to the back. In addition to the central mask, there are often additional, smaller ones. Sometimes the statue depicts the mouth of a predator, sometimes bulls, many so-called solar symbols. They come in different styles, but usually it is a circle inscribed in a square, a kind of mandala, a symbol of the cosmos. This sign is now an official symbol, on the state flag and the state emblem of modern Khakassia. It was discussed that vertical steles might be used as the ancient tool of orientation in space-time milestones and gnomons – sundial of solar hours calendars.” ref
“A graphical drawing of vertical sundial can be seen in the divergent rays on sun-facing stele, where the tooth is a benchmark for the accurate determination of noon. The Okunev culture erected monumental stelae at gravesites. They were either anthropomorphic or zoomorphic with geometric patterns. Steles often incorporated a human head, bent forward slightly. The steles were often re-used by later cultures. For example the Early Turks (Gökturks) often inscribed them with Old-Turkish runic inscriptions, such as the Orkhon inscriptions or Yenisei inscriptions.” ref
“The following artistic features are distinguished:
- free scatter of figures in the pictorial field;
- the presence of anthropomorphic masks;
- elongated proportions of stylized figures;
- a variety of fantastic animals;
- anthropomorphic creatures with bird and animal heads;
- the sacred (world) mountain in the form of a triangle, divided into parts;
- triadic compositions, in which the image of a female deity or its symbol is flanked by two figures of a person or animal;
- images of deities in pointed hats and with bull horns;
- images of Janus anthropomorphic deities;
- images of anthropomorphic figures with two eagle heads;
- images of birds and ornithomorphic figures with a spiral “tuft” on their heads;
- figures of a man with legs and head turned in profile, and the body in front;
- images of characters under the arch of the “firmament”;
- solar sign. ref
“Similarities have been noted between the geometrical anthropomorphic motifs of the Afanasievo culture and Okunev culture of the Minusinsk basin in Siberia, and those on the earlier potteries of Banpo (c. 4000 BCE), of the Yangshao culture in northern China. Pottery styles emerging from the Yangshao culture are known to have spread westward to the Majiayao culture, and then further to Xinjiang and Central Asia. The Okunevo culture, together with the spread of the Seima-Turbino material culture, may be in part be linked to the expansion of Proto-Uralic speakers. Peyrot (2019) argues that “the Okunevo Culture is not to be identified with early Samoyedic, but with Proto-Uralic.” ref
“This is consistent with Janhunen’s convincing arguments that the Ural-Altaic typological profile of Uralic and the primary split between Samoyedic and Finno-Ugric point to an eastern origin (2001; 2009), and it would be just in time for Finno-Ugric to split off and move west towards the Ural Mountains, where this branch was influenced by Proto-Indo-Iranian (e.g. Kuz’mina 2001).” A. G. Kozintsev (2023) argues that the Okunevo culture is better associated with a Yeniseian-related group, possibly Burushaski or an extinct Yeniseian branch. According to him, a Uralic affiliation is unlikely, as Uralic was spoken by people with different material culture, although contact with early Uralic-speakers is plausible. He also reject a possible Indo-Iranian linguistic affiliation, as although the Okunevo culture displays influence from Indo-Iranian groups, they show continuity with previous Ancient Paleo-Siberians, rather than with the Yamnaya culture.” ref

“Abstract: Kurds from Iraq (Dohuk and Erbil Area, North Iraq) have been analyzed for HLA genes. Their HLA genetic profile has been compared with that of other Kurd groups from Iran and Tbilisi (Georgia, Caucasus) and also Worldwide populations. A total of 7,746 HLA chromosomes have been used. Genetic distances, NJ dendrograms and correspondence analyses have been carried out. Haplotype HLA-B*52—DRB1*15 is present in all three analyzed Kurd populations. HLA-A*02-B*51-DRB1*11 is present in Iraq and Georgia Kurds. Haplotypes common to Iran and Iraq Kurds are HLA DRB1*11—DQB1*03, HLA DRB1*03—DQB1*02, and others in a lower frequency. Our HLA study conclusions are that Kurds most probably belong to an ancient Mediterranean / Middle East / Caucasian genetic substratum and that present results and those previously obtained by us in Kurds may be useful for Medicine in future Kurd transplantation programs, HLA Epidemiology (HLA linked diseases) and Pharmacogenomics (HLA-associated drug side effects) and also for Anthropology. It is discussed that one of the most ancient Kurd ancestor groups is in Hurrians (2,000 years BCE).” ref
“HLA is the most polymorphic genetic system described in man. It contains several linked loci which encode for cell surface proteins that have an important function in activating immune response after antigenic presentation. New allele variants are frequently being described (i.e.: 1,883 HLA-DRB1 alleles have been recorded by June 2016). HLA gene frequencies have both a large degree of variability among populations and a striking geographical correlation. These frequencies are useful to infer genetic background and ethnical constitution of modern human groups and also for inferring migrations of ancient ones. In addition, certain combinations of contiguous alleles between HLA neighboring loci show a characteristic frequency due to the robust linkage disequilibrium among them or are distinctive in many extant populations. Also, HLA allele frequencies are unique for studying the origins of relatively homogeneous groups, like the Kurd people living in Iraq.” ref
“On the other hand, Kurd people live in different countries in the Near East such as Syria, Armenia, Turkmenistan, Kazakhstan, Turkey, Iraq and Iran, the so called Kurdistan (“land of Kurds”) (Fig 1, Table 1). Kurdistan is a region placed South Caucasus and North of ancient Mesopotamia. According to genetic studies (like HLA) in Turkish and Kurdish populations, a Anatolian-Mediterranean source for both populations was put forwards; it may be possible that Kurds are initially coming from ancient Hurrians, reviewed in. Studies performed with mtDNA and Y-chr have also been done for Kurds, however there is no firm conclusion to infer that most Kurd people have originated either from Middle East and/or from Central Asia. Most probably, Kurd people gene pool majority may be composed of an admixture of North Mesopotamian (Caucasus) and Near East peoples; Central Asia gene input is not discarded. Kurds have mainly been defined by their ancestry, language, and cultural uses. Estimations of Kurds number are nowadays between 23 to 41 million people; see Table 1 for numbers and country distribution.” ref
“In the present paper, a population of Kurds living in North Iraq (Dohuk and Erbil area, North Mosul, Fig 1) has been studied in order to: 1) Determine the HLA class I (A, B and C) and class II (DRB1 and DQB1) allelic Kurd lineages (hereafter “alleles” for simplicity) and specific HLA haplotypes by using standard DNA based techniques, 2) Compare Iraq Kurd HLA profile with those of Central Asia, Siberian, Mediterranean and other World ethnic groups (Table 2) with specific computer programs in order to find out bases of HLA and disease linkage and origins of Kurd people using genetic distances comparisons, Neighbour Joining (NJ) trees and correspondence analyses, 3) obtaining Kurd HLA profile that may be used for preventive HLA Pharmacogenomics and a virtual regional future transplant waiting list among population, and finally 4) Kurd HLA profiles from Tbilisi (Georgia-Caucasus), Iran and Iraq are also compared among themselves.” ref
“The expected and observed allele frequency values for HLA-A, -B, -C, -DRB1 and -DQB1 shows that the population is in Hardy-Weinberg equilibrium. Table 3 depicts HLA allele frequencies found in the sampled population. Sixteen different HLA-A, twenty-seven different HLA-B and thirteen different HLA-C alleles were founds in class I. Only seven HLA-A alleles, nine HLA-B alleles and seven HLA-C alleles had frequencies higher than 4% (-A*01, -A*02, -A*03, -A*11, -A*24, -A*26, -A*32, -B*07, -B*08, -B*18, -B*35, -B*38, -B*41, -B*44, -B*51, -B*52, -C*04, -C*06, -C*07, -C*12, -C*15, -C*16 and -C*17). Twelve different HLA-DRB1 alleles and five different HLA-DQB1 alleles were found. Only eight HLA-DRB1 alleles and four HLA-DQB1 alleles had frequencies higher than 4% (-DRB1*01, -DRB1*03, -DRB1*04, -DRB1*07, -DRB1*11, -DRB1*13, -DRB1*14, -DRB1*15, -DQB1*02, -DQB1*03, -DQB1*05 and -DQB1*06).” ref
“DRB1 alleles were used to compare our three Kurd samples with other populations in NJ analysis. It was not possible to perform this study with HLA class I allele frequencies due to the lack of class I studies in many worldwide available populations (Table 2). NJ relatedness dendrogram based on HLA-DRB1 analysis separates populations in two differentiated clusters: A and B (Fig 2). Cluster A groups North and South Mediterraneans (Europeans and Africans) and Middle East populations (included Iraq Kurds, Iran Kurds, and Georgia Kurds). Cluster B includes Central and eastern Siberians and Oriental population: such Tuvinians, Todja, Tofalar, Ulchi, Negidal, Japanese, Kets, Evenks. Cluster A1 contains Iraq Kurds and Georgia Kurds, which are placed together, Palestinians, Armenians, Iran Kurds, East Europe (Croatians), Cretans, Macedonians, Near East (Lebanese), and Non-Ashkenazi Jews(Fig 2). Cluster A2 places together other Mediterraneans (Europeans and Africans such as Spaniards and Spanish Basques), French, Berbers, and Moroccans (Fig 2).” ref
“Correspondence analysis based on HLA-DRB1 allele frequencies (Fig 3) shows similar results to those of Fig 2. Two clusters are clearly defined according to first dimension that explains most of the variability among populations. The first one groups together Siberian and Oriental populations (left, Fig 3), and the second cluster comprises Europeans, Mediterraneans, Caucasus, and Iranian populations; Iraq Kurds, Iran Kurds, and Georgia Kurds are located relatively close together. Plain genetic distances (DA) show that Iraq Kurds’ closest genetic distances are the following: Near East populations (Iran Kurds, Palestinians, FarsParsi, Georgia Kurds, and Ashkenazi Jews), eastern Mediterranean populations (Armenians, Cretans, and Macedonians), and Mediterranean populations (Sardinians, Spaniards, Algerians, and Italians).” ref
HLA-A, -B. DRB1 and -DQB1 extended haplotype analysis in Kurds: comparison with other populations
“Associations between different HLA loci were estimated in Iraq Kurds (Table 4). The most frequent five loci haplotype (A-B-C-DRB1-DQB1) were obtained. Eight Mediterranean five loci haplotypes are found (A*03-B*44-C*16-DR*04-DQ*03, A*26-B*08-C*07-DR*03-DQ*02, A*33-B*14-C*08-DR*01-DQ*05, A*01-B*52-C*12-DR*15-DQ*06, A*01-B*08-C*07-DR*03-DQ*02, A*02-B*44-C*05-DR*11-DQ*03, A*01-B*35-C*04-DR*14-DQ*05, and A*02-B*51-C*16-DR*15-DQ*06) and three Near East five loci haplotypes (A*03-B*35-C*04-DR*11-DQ*03, A*02-B*51-C*15-DR*11-DQ*03 and A*02-B*51-C*14-DR*11-DQ*03) represents about 4.01% (Table 4 and footnote). Also, an Eurasiatic haplotype is found: A*24-B*35-C*04-DR*11-DQ*03. These results show HLA genetic characteristics of both Mediterranean and Near East populations; also, one Eurasiatic haplotype is found in a relatively high frequency.” ref
HLA haplotype common to Georgian, Iran, and Iraq Kurds
“The following haplotypes are shared among Kurd populations: HLA-B*52-DRB1*15 is present in Iran Kurds (2.5%), Iraq Kurds (1.59%), and Georgia Kurds (3.6%); these later results have been taken from our previous work with a low number of individuals (n = 30). HLA-A*02-B*51-DRB1*11 is present in Iraq Kurds (2.3%) and Georgia Kurds (3.6%); DRB1*11-DQB1*03 is present in Iraq Kurds (10%) and Iran Kurds (6.7%); DRB1*03-DQB1*02 is present in Iraq Kurds (5.9%) and Iran Kurds (3.38%); DRB1*01-DQB1*05 is present in Iraq Kurds (3.3%), and Iran Kurds (1.67%) and DRB1*15-DQB1*06 is present in Iraq Kurds (2.5%) and in Iran Kurds (1.59%).” ref
“Kurds are currently living in Kurdistan, a region encompassing different parts of several Middle East countries (Fig 1, Table 1); in addition they have moved to live in Middle East and European cities. We had previously studied Kurds in Tbilisi (Georgia) and also in Iran for HLA allele frequencies. Our conclusions were that their HLA profile showed that Kurds form part of Mediterranean stock of people and also had Caucasus genetic traits (Svan, Georgians). Also, it is worth mentioning that Lak population (East Caucasus Area) may be close to both Lur and Kurd populations, and Lak name could be considered composed of Lur and Kurd words. In the present paper, we have analyzed HLA genes in Kurds living in North Iraq (Erbil and Dohuk areas). Comparison with other populations place them as a Middle East population, close to Kurds from Tbilisi-Georgia, Palestinians, Armenians and Kurds from Iran (Fig 2).” ref
“Correspondence analysis (Fig 3, right side) shows that Kurds (living in Iran, Tbilisi-Georgia and Iraq) somewhat divide bidimensional representation analysis in western Mediterraneans (upper part) and Eastern Mediterraneans (lower part). In both analyses (Figs (Figs22 and and3),3), Kurds are also close to Caucasian (Svan, Georgian) populations. Conclusion is that Kurds are genetically close to surrounding Caucasian and Mediterranean populations and that have remained settled down in Kurdistan since ancient times; supporting historical evidence is reviewed in our previous work. HLA genetic similarity have been reported between Turks (whose genes belong to old Anatolian stock) and Kurds. Kurds and Turks speak languages that are included in different families. However, Kurd HLA genetic studies include them into Mediterranean stock together with Turks.” ref
“Other genetic studies based on Y-Chr in Kurds from Turkey, Georgia, and Iran identify the dominant presence of haplogroups originated in the Middle East (Anatolia or Mesopotamia) that show a close association with Jews, Lebanese, and Turkish genes. Also, Iranian populations are close to Kurds. This again shows that languages and genes do not correlate because languages may be imposed by a genetic (but powerful) minority. This is the case of Turks: Anatolian people were settled down there since ancient prehistoric times, but a minority of people (Turks) coming from Central Asia imposed language in historical times.” ref
“Thus, Middle Eastern peoples from the Mediterranean border and Kurds originally seem to belong to a similar ethnic group according to the results of HLA autonomic and Y chromosome genes. Kurds have always lived in the mountains being “autochthonous” (6000 BCE). Hurrians, whose language was Caucasian (and not Indo-European) may be Kurds ancient genetic background. By 1200 BCE, Medes and others invaded Hurrian area. Kurdish historians consider that Kurds come from Medes. “Kuru” was the first name of Kurds given by Assyrians (1000 BCE) to groups living at Mt. Azu, Kurdistan. Kurds are also mentioned by early classical historians like Polybius (133 BCE) and Strabo (48 CE). Kurds were named “the Mountains People” under Persian, Greek, and Roman Anatolian Peninsula rule.” ref
“In summary, all three Kurd populations studied in the present paper are genetically close together and to other Mediterranean and Caucasus populations according to HLA genes. This study may also help for future transplantation programs in their area and Kurd HLA Epidemiology and Pharmacogenomics.” ref

The Genomic History of the Bronze Age Southern Levant
“Researchers report genome-wide DNA data for 73 individuals from five archaeological sites across the Bronze and Iron Ages Southern Levant. These individuals, who share the “Canaanite” material culture, can be modeled as descending from two sources: (1) earlier local Neolithic populations, and (2) populations related to the Chalcolithic Zagros or the Bronze Age Caucasus. The non-local contribution increased over time, as evinced by three outliers who can be modeled as descendants of recent migrants. We show evidence that different “Canaanite” groups genetically resemble each other more than other populations. We find that Levant-related modern populations typically have substantial ancestry coming from populations related to the Chalcolithic Zagros and the Bronze Age Southern Levant. These groups also harbor ancestry from sources we cannot fully model with the available data, highlighting the critical role of post-Bronze Age migrations into the region over the past 3000 years. Genome-wide data from Bronze Age individuals across nine sites in the Southern Levant show strong genetic resemblance including a component from populations related to Chalcolithic Zagros and Early Bronze Age Caucasus introduced by gene flow lasting at least until the late Bronze Age and impacting modern Levantine population architecture.” ref
“The Bronze Age (ca. 3500–1150 BCE) was a formative period in the Southern Levant, a region that includes present-day Israel, Jordan, Lebanon, the Palestinian Authority, and southwest Syria. This era, which ended in a large-scale civilization collapse across this region (Cline, 2014), shaped later periods both demographically and culturally. The following Iron Age (ca. 1150–586 BCE) saw the rise of territorial kingdoms such as biblical Israel, Judah, Ammon, Moab, and Aram Damascus, as well as the Phoenician city-states. In much of the Late Bronze Age, the region was ruled by imperial Egypt, while in later phases of the Iron Age, it was controlled by the Mesopotamian-centered empires of Assyria and Babylonia. Archaeological and historical research has documented major changes during the Bronze and Iron Ages, such as the cultural influence of the northern (Caucasian) populations related to the Kura-Araxes tradition during the Early Bronze Age (Greenberg and Goren, 2009) and impacts from the “Sea Peoples” (such as Philistines) from the west in the beginning of the Iron Age (Yasur-Landau, 2010).” ref
“The inhabitants of the Southern Levant in the Bronze Age are commonly described as “Canaanites”, that is, residents of the Land of Canaan. The term appears in several 2nd millennium BCE sources (e.g., Amarna, Alalakh, and Ugarit tablets) and in biblical texts dating to the 8th-7th centuries BCE and later (Bienkowski, 1999; Lemche, 1991; Na’aman, 1994a). In the latter, the Canaanites are referred to as the pre-Israelite inhabitants of the land (Na’aman, 1994a). Canaan of the 2nd millennium BCE was organized in a system of city-states (Goren et al., 2004), with elites ruling from urban hubs over rural (and in some places pastoral) countryside. The material culture of these city-states was relatively uniform (Mazar, 1992), but whether this uniformity extends to their genetic ancestry is unknown. Whereas genetic ancestry and material culture are unlikely to ever match perfectly, past ancient DNA analyses show that they might sometimes be strongly associated. In other cases, a direct correspondence between genetics and culture cannot be established. We discuss several examples in the Discussion.” ref
“Previous ancient DNA studies published genome-scale data for 13 individuals from four Bronze Age sites in the Southern Levant: three individuals from ‘Ain Ghazal in present-day Jordan, dated to ~2300 BCE (Intermediate Bronze Age) (Lazaridis et al., 2016); five from Sidon in present-day Lebanon, dated to ~1750 BCE (Middle Bronze Age) (Haber et al., 2017); two from Tel Shadud in present-day Israel, dated to ~1250 BCE (Late Bronze Age) (van den Brink et al., 2017); and three from Ashkelon in present-day Israel, dated to ~1650–1200 BCE (Middle and Late Bronze Age) (Feldman et al., 2019). The ancestry of these individuals could be modeled as a mixture of earlier local groups and groups related to the Chalcolithic people of the Zagros Mountains, located in present-day Iran and designated in previous studies as Iran_ChL (Haber et al., 2017; Lazaridis et al., 2016). The Bronze Age Sidon group could be modeled as a major (93±2%) ancestral source for present-day groups in the region (Haber et al., 2017).” ref
“A study of Chalcolithic individuals from Peqi’in cave in the Galilee (present-day Israel) showed that the ancestry of this earlier group included an additional component related to earlier Anatolian farmers, which was excluded as a substantial source for later Bronze Age groups from the Southern Levant, with the exception of the coastal groups from Sidon and Ashkelon (Feldman et al., 2019; Harney et al., 2018). These observations point to a degree of population turnover in the Chalcolithic-Bronze Age transition, consistent with archaeological evidence for a disruption between local Chalcolithic and Early Bronze cultures (de Miroschedji, 2014). Here, we set out to address three issues. First, we sought to determine the extent of genetic homogeneity among the sites associated with Canaanite material culture. Second, we analyzed the data to gain insights into the timing, extent, and origin of gene flow that brought Zagros and Caucasus-related ancestry to the Bronze Age Southern Levant. Third, we assessed the extent to which additional gene flow events have impacted the region since that time.” ref
“To address these questions, we generated genome-wide ancient DNA data for 71 Bronze Age and two Iron Age individuals, spanning roughly 1,500 years, from the Intermediate Bronze Age to the Early Iron Age. Combined with previously published data on the Bronze and Iron Ages in the Southern Levant, we assembled a dataset of 93 individuals from nine sites across present-day Israel, Jordan, and Lebanon, all demonstrating Canaanite material culture. We show that the sampled individuals from the different sites are usually genetically similar, albeit with subtle but, in some cases, significant differences, especially in residents of the coastal regions of Sidon and Ashkelon. Almost all individuals can be modeled as a mixture of local earlier Neolithic populations and populations from the northeastern part of the Near East. However, the mixture proportions change over time, revealing the demographic dynamics of the Southern Levant during the Bronze Age. Finally, we show that the genomes of present-day groups geographically and historically linked to the Bronze Age Levant, including the great majority of present-day Jewish groups and Levantine Arabic-speaking groups, are consistent with having 50% or more of their ancestry from people related to groups who lived in the Bronze Age Levant and the Chalcolithic Zagros. These present-day groups also show ancestries that cannot be modeled by the available ancient DNA data, highlighting the importance of additional major genetic impacts on the region since the Bronze Age.” ref
“Abstract: Despite the localisation of the southern Caucasus at the outskirt of the Fertile Crescent, the Neolithisation process started there only at the beginning of the sixth millennium with the Shomutepe-Shulaveri culture of yet unclear origins. We present here genomic data for three new individuals from Mentesh Tepe in Azerbaijan, dating back to the beginnings of the Shomutepe-Shulaveri culture. Reserchers evidence that two juveniles, buried embracing each other, were brothers. We show that the Mentesh Tepe Neolithic population is the product of a recent gene flow between the Anatolian farmer-related population and the Caucasus/Iranian population, demonstrating that population admixture was at the core of the development of agriculture in the South Caucasus. By comparing Bronze Age individuals from the South Caucasus with Neolithic individuals from the same region, including Mentesh Tepe, we evidence that gene flows between Pontic Steppe populations and Mentesh Tepe-related groups contributed to the makeup of the Late Bronze Age and modern Caucasian populations. Our results show that the high cultural diversity during the Neolithic period of the South Caucasus deserves close genetic analysis.” ref
“This article presents the results of analyses carried out by several specialists on some of the raw materials (metal, obsidian, and semi-precious stones) found at Mentesh Tepe (middle Kura Valley, Azerbaijan) and dating from the Late Chalcolithic and Early Bronze Age periods. They show that the supply area is located at a distance of 30 to 300 km from the site. The sources of copper and arsenic used during the Late Chalcolithic are found in the “volcanogenic massive sulfide” (VMS) deposits that stretch to the south of the site in the Lesser Caucasus. The question of their origin during the Early Bronze Age remains open, as does that of gold and tin at the same time. The provenance analyses of the obsidian show that several sources were used, some of which, located in present-day Turkey, were very distant. Gegham, Armenia, was by far the most important supply area in the Late Chalcolithic, while Chikiani, Georgia, may have played this role in the Early Bronze Age. Finally, the raw materials used for beads come from ophiolitic strata and volcanic contexts in the Lesser Caucasus located not far from Mentesh Tepe.” ref
“The Chalcolithic or Copper Age is the transitional period between the Neolithic and the Bronze Age. It is taken to begin around the mid-5th millennium BCE, and ends with the beginning of the Bronze Age proper, in the late 4th to 3rd millennium BCE, depending on the region.” ref
“During the Late Chalcolithic period, in the second half of the 5th millennium (between 4,360 and 4,115 in median radiocarbon dates), remains of one or several buildings made of unbaked mud-bricks as well as courtyards have been brought to light. Excavations have shown that different craftsmanship activities took place both inside and outside the buildings, including metallurgy, lithic, and pottery production. Several features of this period have been interpreted as related with northern Mesopotamia.” ref
“During the Early Bronze Age, the place seems to have had only a funerary character with the implantation of two successive kurgans with multiple burials and several individual burials. A great number of pits have also been related to this period. The first kurgan (ST 4) is linked with the beginning of the Kura-Araxes culture and dated to the end of the 4th millennium (between 3,155 and 2,986 in median radiocarbon dates); it contained at least 39 individuals but gave only little funerary material – among which mainly stone beads – and was ritually destroyed by fire at the end of its use (Lyonnet et al. 2015). The second kurgan (ST 54) is related to the Martkopi phase of the Early Kurgan culture/end of the Kura-Araxes culture and dated between 2495 and 2408 in median radiocarbon dates; it contained three individuals and was fairly rich in jewelry (metal and stone) (Pecqueur, Decaix, Lyonnet 2017).” ref
“One among the most interesting discoveries made at Mentesh Tepe is the number of finds linked with metallurgy during the Late Chalcolithic period. Several articles have already been published (for instance Courcier 2012; Courcier forthcoming). Very few other sites dating to this period – except Değirmentepe in eastern Turkey, unfortunately inadequately published, and Ovçular Tepesi in Nakhchivan (Gailhard et al. 2017) – have yielded such abundant data on local metallurgy. Beside a great number of objects, mainly awls and rings, discovered in different places in and outside the buildings, all the different steps of the operational chain leading to them, have been brought to light: ores, crucibles, slags, molds and ingots (Courcier forthcoming). Correcting an earlier theory (Chernykh 1992), this important discovery places the southern Caucasus on a level comparable with that of the Balkan “province” where metallurgy was already carried on during the second half of the 5th millennium BCE.” ref
“Most of the manufactured objects are made from unalloyed copper with significant traces of lead and silver, but 13 are in arsenical copper (1.1‑3.2%) (Courcier forthcoming). Archaeometallurgical studies have been made, including mineralogical investigations (thin‑section, SEM‑EDS, XRD, metallographies) and chemical ones (ICP‑MS) at the Bochum Laboratory of the Deutsches Bergbau Museum under the direction of A. Hauptmann and M. Prange , as well as isotopic analyzes (HR‑MC‑ICP‑MS) at the department of petrology and geochemistry of the Goethe University in Frankfurt under the direction of Prof. Dr Sabine Klein. The aim was to identify the possible sources for the metal ores. They have shown two possible areas for these procurements, west and south of Mentesh Tepe. The first possible one (west) includes three metallogenic zones: Bolnissi-Madneuli (Madneuli deposit), north of Alaverdi Kapan (Shamlug, Akhtala, Agvi, Ankadzor deposits) and north of Pambak-Zangezur (Fioletovo deposit). The second one (south) includes two metallogenic zones: Sotk-Gosha (Gosha) and Kedabek (Siny Yar, Kedabek).” ref
“These results point at three possible areas within the “volcanogenic massive sulfide deposits” (VMS) for the origin of the copper ores: eastern Armenia (districts of Alaverdi and Vanadzor), southern Georgia (district of Madneuli), or south-west Azerbaijan ( districts of Gosha and Kedebek), ie in a radius varying between 40 and 180 km from Mentesh Tepe. The Early Bronze Age period at Mentesh Tepe is void of traces of architecture except for two kurgans, several individual graves, and a large number of pits where the material is mixed with that of the earlier periods and therefore difficult to date with certainty. We will then only deal here with the metal found in secure contexts.” ref
“At the beginning of the Early Bronze Age, in kurgan ST 4, only a few items have been found, made either of fired clay (21 pots) or of stone (hundreds of beads, see below), but metal artifacts were singularly absent, possibly because they had been collected before the kurgan was set on fire for ritual reasons (Lyonnet et al. 2015). The metal items we have from secure contexts come essentially from the second kurgan, ST 54, which is related to the Martkopi phase of the Early Kurgan Culture (or the end of the Kura-Araxes culture) (Pecqueur, Decaix, Lyonnet 2017). These items consist of spiral-shaped hair rings found near the skulls of two women skeletons. One of the women also wore two spiral bracelets and, probably sewn on her costume near the shoulder, a small umbo or mastos-shaped casket.” ref
“Analyzes by ICP-MS have shown that the spiral-shaped hair-rings and the bracelets are made of bronze (from 2 to 11% Sn), with small amounts of arsenic (0.15 to 0.45% As) and significant traces of lead, while the umbolum-shaped casket is made of a silver-copper alloy (49.8% Ag, 42.9% Cu). We still have no clue as to the provenance of tin, but tin is rare in the Caucasus so that it probably comes from elsewhere. The provenance of the copper is still under question: either Alaverdi-Kapan or Gosha in the Lesser Caucasus, or with a foreign origin, like the tin. Several gold beads, a gold hair-spiral and a gold ring have also been discovered in this kurgan and are related to a third person, an aged man. At first, it seemed plausible that the gold came from Sakdrisi, an important gold mine in the Madneuli district of Georgia exploited during the Early Bronze Age (Gambashidze, Stöllner 2016), but the analysis made by M. Jansen at the Bochum Laboratory and at the Institute for Geosciences of the Goethe University (Frankfurt) has demonstrated that this is not the case.” ref
“For both the ring and a bead, observation with a digital microscope has shown the presence of inclusions made of platinum group elements (PGE), i.e., natural alloys of osmium, iridium, and ruthenium (Jansen et al., this volume). These cannot be found in primary gold deposits such as Sakdrisi, but they derive from chromite in ultrabasic rocks. These rocks eroded, and the PGE minerals have accumulated together with gold from a primary gold source in a secondary placer deposit. The PGE minerals were subsequently incorporated into the artifacts after panning and melting of the gold from this deposit. A XRF analysis has shown that the bead is an alloy mainly consisting of gold and silver, with some copper ( tab. 1 ). Both the ring and the bead correlate together well in their tin and platinum content with most of the artifacts from the Lesser Caucasus that have been analyzed (Jansen et al., this volume). This, together with the presence of PGE inclusions, shows that the metal used is placer gold, ie from alluvial origin, but its exact provenance cannot be established for the moment.” ref
“A large number of obsidian tools and flakes and a few nuclei have been discovered in all the levels of occupation at the site. The obsidian items present a wide range of colors and appearances. The technology used for the tool production is still under study by L. Astruc, D. Guilbeau, and A. Samzun (for preliminary studies, see Astruc, Samzun, Gratuze 2012; Guilbeau, Astruc, Samzun 2017). In order to trace the origin of the raw material, a large number of samples have been analyzed. In addition, samples of obsidian fragments used as a temper for some Late Chalcolithic pottery types – essentially cooking pots – have also been analyzed (Palumbi et al. 2018).” ref
“For the Chalcolithic period, the studied sample includes a total of 268 lithic elements (blade production and flakes) coming from well stratified contexts and 78 inclusions from 47 ceramic pots. We have less data for the Kura-Araxes chipped stone as this period is mainly represented on the site by two kurgans and pits, the context of which is often mixed with earlier material. Nevertheless, on a technological basis, four artifacts have been chosen and characterized by geochemistry. Altogether, this represents, so far, the largest archaeological sample that has been geochemically characterized among the Caucasian and Near Eastern sites.” ref
“The Chalcolithic is part of prehistory, but based on archaeological evidence, the emergence of the first state societies can be inferred, notably in the Fertile Crescent (notably Sumer) Predynastic Egypt, and Proto-Minoan Crete, with late Neolithic societies of comparable complexity emerging in the Indus Valley (Mehrgarh), China, and along the north-western shores of the Black Sea. The development of states—large-scale, populous, politically centralized, and socially stratified polities/societies governed by powerful rulers—marks one of the major milestones in the evolution of human societies. Archaeologists often distinguish between primary (or pristine) states and secondary states. Primary states evolved independently through largely internal developmental processes rather than through the influence of any other pre-existing state.” ref
“The earliest known primary states appeared in Anatolia c. 5200 BCE, in Mesopotamia c. 3700 BCE, in Greece c. 3500 BCE, in Egypt c. 3300 BCE, in the Indus Valley c. 3300 BCE, and in China c. 1600 BCE. In Africa, discoveries in the Agadez Region of Niger evidence signs of copper metallurgy as early as 2000 BCE. This date pre-dates the use of iron by a thousand years.Copper metallurgy seems to have been an indigenous invention in this area, because there is no clear evidence of influences from Northern Africa, and the Saharan wet phase was coming to an end, hindering human interactions across the Saharan region. It appeared to not be fully developed copper metallurgy, which suggests it was not from external origins.” ref
| City | period |
|---|---|
| Anau | 4000 to 1000 BCE |
| Anshan | 4000 to 1000 BCE |
| Bad-tibira | 5000 to 2300 BCE |
| Çatalhöyük | 6700 to 5700 BCE |
| Güvercinkayası | 5200 to 4750 BCE |
| Ebla | 3500 to 1600 BCE |
| Eridu | 5400 to 2050 BCE |
| Girsu | 5000 to 2100 BCE |
| Heliopolis (Lower Egypt) | 3500 to ~1000BCE |
| Isin | 3500 to 2100 BCE |
| Jericho | 9600 to 1400 BCE |
| Kish | 4000 to 2300 BCE |
| Knossos | 7000 to 1900 BCE |
| Lagash | 4000 to 2250 BCE |
| Laish | 4500 to 1350 BCE |
| Mari | 2900 to 1759 BCE |
| Mehrgarh | 5500 to 2500 BCE |
| Nekhen (Upper Egypt) | 3500 to ~150BCE |
| Nippur | 5000 to 2450 BCE |
| Susa | 4200 to 2330 BCE |
| Rakhigarhi | 6500 to 1900 BCE |
| Ugarit | 6000 to 1190 BCE |
| Ur | 4000 to 2000 BCE |
| Uruk | 4000 to 3100 BCE |
Mentesh Tepe, an early settlement of the Shomu-Shulaveri Culture in Azerbaijan
“Abstract: Excavations at Mentesh Tepe, western Azerbaijan, have unearthed Neolithic levels dated to an early stage of the Shomu-Shulaveri Culture, with a specific material culture and several inhumations among which a multiple burial. At that stage, already a full domestication of plants and animals is evident. Many questions have been raised concerning the origins of this culture, and its end is also still obscure. Relations with societies in the north-Mesopotamian area have again recently been evidenced at its beginnings. Mentesh Tepe, with its exceptional succession of occupations from the Neolithic to the Early Bronze Age, could help providing some clues for the links between the Neolithic and the Chalcolithic periods. The site is presented here under different points of views (architecture, burials, material culture) but in a preliminary stage since many studies are still in progress. Questions are raised about the climate and the apparent absence of pre- and post-Shomu-Shulaveri Culture possibly due to silting or erosion processes linked with the mobility of the Caspian Sea level.” ref
“The Shomu-Shulaveri Culture (hereafter SSC) has first been identified about half a century ago by I. Narimanov, during excavations done in Western Azerbaijan on the sites of Shomu Tepe, Tojre-Tepe and Babadervish, and later on other sites in the vicinity. Further discoveries were made by A.I. Dzhavakhishvili and T.N. Chubinishvili a few years later in Eastern Georgia at Shulaveris Gora and other sites around where similar architecture and material culture were unveiled. These and subsequent excavations have shown that several clusters of settlements sharing the same cultural features spread on the northern foothills of the Lesser Caucasus along a series of small tributaries of the Kura River in the middle of its course, from the Khramis in the North to the Zeyem Chaj in the South, astride actual Georgia and Azerbaijan.” ref
“A typical circular architecture with a wealth of material (pottery, bone-, obsidian- and stone tools) was retrieved from these sites together with rare discoveries of metal items. Specific studies were made on the part of the tools that underlined their main use for agriculture or on animal skins. Attempts were made to organize the material from the excavations in 5 successive phases rising in complexity. Most of the plant remains (wheat, barley, pips of grape) and animal bones (caprines, bovids, pigs, and dogs) pointed at an already advanced stage of domestication where caprines played a leading role. This led to consider that this culture was that of sedentary groups already well akin with farming and breeding, and, due to the presence of a few metal objects, it was dated to the Eneolithic/Chalcolithic period.” ref
“Contradictory hypotheses were raised about the climate. On the one hand, some considered that it was similar to nowadays or possibly drier on the basis of pollen analysis from two sites, Imiris Gora and Arukhlo 1. They also thought that the birth of agriculture was in itself evidence that climate was getting more arid since wild cereals were not sufficient anymore. On the other hand, others proposed that it was more humid than today, as sites were generally positioned along old, and now dried, courses of the rivers. Other sites dated to the same “Eneolithic/Coper age” period were discovered at about the same time in south-east of Azerbaijan during surveys and soundings in the Mil’ and Karabakh steppes, and two
sites were excavated (Alikemek in the Mugan steppes, and Kül Tepe in Nakhichevan). A complete Halaf pot, compared to those of Tilki Tepe near Van, had been found in the lowest levels of Kül Tepe, and probably comes from a grave, though this is not clearly mentioned. While some authors considered these SE sites as belonging to a different cultural group, mainly on the basis of the presence of painted pottery, others proposed that they and the SSC ones were local variants of one unique culture because of similarities in the lithic and bone industries.” ref
“Many questions were raised as to the origins of this/these cultures, since previous “Neolithic” remains in these territories from which they could have raised were unknown at that time, except for very few and doubtful cases. Only Western Georgia had caves and stations dating to an older period, but no links could be established with the central and eastern part of Transcaucasia. General similarities in the architecture and in part of the material of the SSC and SE sites with that of cultures situated further south was mentioned. Some authors considered that the cultures of northern Mesopotamia, especially the Hassuna and Halaf communities, had probably played an important role in the formation of the SSC, while those of northern Iran and the Zagros had influenced the painted pottery of the SE group.” ref
“Altogether, many uncertainties prevented a clear understanding of both the SSC and the SE cultures, among which stood the problems of their absolute dates and a better definition of their features. New calibrated dates and further research have since then established with certainty that the SSC belongs to the Late Neolithic and that it developed along the 6th millennium. Several new excavations are now going on, and numerous analyses are being made that intend to understand the origins, development, economy, and environment of the SE cultures in the Mil Plain and, even more research is going on the SSC, at Aruchlo, at G€ oy Tepe, at HacıElamxanlı Tepe, and at Gadachrili Gora (Hamon et al., this volume).” ref
“Sites outside of the Kura Valley, like Aratashen and Aknashen/Khaturnakh in the Ararat Plain of Armenia, are also associated to the SSC due to similarities in their architecture and material culture. Finally, recent discoveries made at Mentesh Tepe can now be added to this list. The aim of this article is to present some of the preliminary results of the 2011e2014 excavations which took place at Mentesh and which place it among the earliest known settlements of the SSC. This corrects what had first been proposed, i.e. an early Chalcolithic (Copper age) period, and a Neolithic period for phase 1, followed by an early Chalcolithic period for phase 2.” ref
“Abstract: Archaeogenetic studies have described the formation of Eurasian ‘steppe ancestry’ as a mixture of Eastern and Caucasus hunter-gatherers. However, it remains unclear when and where this ancestry arose and whether it was related to a horizon of cultural innovations in the 4th millennium BCE that subsequently facilitated the advance of pastoral societies in Eurasia. Here we generated genome-wide SNP data from 45 prehistoric individuals along a 3000-year temporal transect in the North Caucasus. We observe a genetic separation between the groups of the Caucasus and those of the adjacent steppe. The northern Caucasus groups are genetically similar to contemporaneous populations south of it, suggesting human movement across the mountain range during the Bronze Age. The steppe groups from Yamnaya and subsequent pastoralist cultures show evidence for previously undetected farmer-related ancestry from different contact zones, while Steppe Maykop individuals harbor additional Upper Palaeolithic Siberian and Native American related ancestry.” ref
“The 1100-kilometre-long Caucasus mountain ranges extend between the Black Sea and the Caspian Sea and are bounded by the rivers Kuban and Terek in the north and the Kura and Araxes rivers in the south (Fig. 1). The rich archaeological record suggests extensive human occupation since the Upper Palaeolithic. A Neolithic lifestyle based on food production began in the Caucasus after 6.000 cal BCE. As a region rich in natural resources such as ores, pastures, and timber, the Caucasus gained increasing importance to the economies of the growing urban centers in northern Mesopotamia. In the 4th millennium BCE, the archaeological record attests to the presence of the Maykop and Kura-Araxes, two major cultural complexes of the Bronze Age (BA) in the region. The Maykop culture is well known for its large and rich burial mounds, especially at the eponymous Maykop site, which reflects the rise of a new system of social organization, while the Kura-Araxes is found on both flanks of the Caucasus mountain range, demonstrating a connection between north and south.” ref
“Contact between the Near East, the Caucasus, the Steppe, and central Europe is documented, both archaeologically and genetically, as early as the 5th millennium BC. This increased in the 4th millennium BCE along with the development of new technologies such as the wheel and wagon, copper alloys, new weaponry, and new breeds of domestic sheep. Such contact was critical in the cultural and genetic formation of the Yamnaya complex on the Eurasian Steppe—with about half of BA Steppe ancestry thought to derive from the Caucasus. In the 3rd millennium BC, increased mobility associated with wheeled transport and the intensification of pastoralist practices led to dramatic expansions of populations closely related to the Yamnaya, accompanied by the domestication of horses allowing more efficient keeping of larger herds. These expansions ultimately contributed a substantial fraction to the ancestry of present-day Europe and South Asia. Thus, the Caucasus region played a crucial role in the prehistory and formation of Eurasian genetic diversity.” ref
“Recent ancient DNA studies have resolved several long-standing questions regarding cultural and population transformations in prehistory. One important feature is a cline of European hunter-gatherer (HG) ancestry that runs roughly from West to East (hence WHG and EHG). This ancestry differs from that of Early European farmers, who are more closely related to farmers of northwest Anatolia and also to pre-farming Levantine individuals. The near East and Anatolia have long-been seen as the regions from which European farming and animal husbandry emerged. In the Mesolithic and Early Neolithic, these regions harbored three divergent populations, with Anatolian and Levantine ancestry in the west and a group with a distinct ancestry in the east. The latter was first described in Upper Pleistocene individuals from Georgia (Caucasus hunter-gatherers; CHG) and then in Mesolithic and Neolithic individuals from Iran. The following millennia, spanning the Neolithic to BA, saw admixture between these ancestral groups, leading to a pattern of genetic homogenization of the source populations. North of the Caucasus, Eneolithic, and BA individuals from the Samara region (5200–4000 BCE) carry an equal mixture of EHG- and CHG/Iranian ancestry, so-called ‘steppe ancestry’ that eventually spread further west, where it contributed substantially to present-day Europeans, and east to the Altai region as well as to South Asia.” ref
“To understand and characterize the genetic variation of Caucasian populations, present-day groups from various geographic, cultural/ethnic, and linguistic backgrounds have been analyzed previously. Yunusbayev and colleagues described the Caucasus region as an asymmetric semipermeable barrier based on a higher genetic affinity of southern Caucasus groups to Anatolian and near Eastern populations and a genetic discontinuity between these and populations of the North Caucasus and the adjacent Eurasian steppes. While autosomal and mitochondrial DNA data appear relatively homogeneous across the entire Caucasus, the Y-chromosome diversity reveals a deeper genetic structure attesting to several male founder effects, with striking correspondence to geography, ethnic and linguistic groups, and historical events.” ref
“In our study, we aimed to investigate when and how the genetic patterns observed today were formed and test whether they have been present since prehistoric times by generating time-stamped human genome-wide data. We were also interested in characterizing the role of the Caucasus as a conduit for gene-flow in the past and in shaping the cultural and genetic makeup of the wider region. This has important implications for understanding the means by which Europe, the Eurasian steppe zone, and the earliest urban centers in the Near East were connected. We aimed to genetically characterize individuals from cultural complexes such as the Maykop and Kura-Araxes and assessing the amount of gene flow in the Caucasus during times when the exploitation of resources of the steppe environment intensified, since this was potentially triggered by the cultural and technological innovations of the Late Chalcolithic and EBA around 4000–3000 BCE.” ref
“Finally, since the spread of steppe ancestry into central Europe and the eastern steppes during the early 3rd millennium BCE was a striking migratory event in human prehistory, we also retraced the formation of the steppe ancestry profile and tested for influences from neighboring farming groups to the west or early urbanization centers further south. Here we show that individuals from our Caucasian time transect form two distinct genetic clusters that were stable over 3000 years and correspond with eco-geographic zones of the steppe and mountain regions. This finding is different from the situation today, where the Caucasus mountains separate northern from southern Caucasus populations. However, during the early BA we also observe subtle gene flow from the Caucasus as well as the eastern European farming groups into the steppe region, which predates the massive expansion of the steppe pastoralists that followed in the 3rd millennium BCE.” ref
“We report genome-wide data at a targeted set of 1.2 million single nucleotide polymorphisms (SNPs) for 59 Eneolithic and BA individuals from the Caucasus region. After filtering out 14 individuals that were first-degree relatives or showed evidence of contamination we retained 45 individuals for downstream analyses using a cutoff of 30,000 SNPs. We merged our newly generated samples with previously published ancient and modern data. We first performed principal component analysis (PCA) and ADMIXTURE analysis to assess the genetic affinities of the ancient individuals qualitatively (Fig. 2). Based on PCA and ADMIXTURE plots, reserchers observe two distinct genetic clusters: one falls with previously published ancient individuals from the West Eurasian steppe (hence termed ‘Steppe’), and the second clusters with present-day southern Caucasian populations and ancient BA individuals from today’s Armenia (henceforth called ‘Caucasus’), while a few individuals take on intermediate positions between the two. The stark distinction seen in our temporal transect is also visible in the Y-chromosome haplogroup distribution, with R1/R1b1 and Q1a2 types in the Steppe and L, J, and G2 types in the Caucasus cluster. In contrast, the mitochondrial haplogroup distribution is more diverse and similar in both groups.” ref
“In contrast, the oldest individuals from the northern mountain flank itself, which are three first-degree-related individuals from the Unakozovskaya cave associated with the Darkveti-Meshoko Eneolithic culture (analysis label ‘Eneolithic Caucasus’), show mixed ancestry mostly derived from sources related to the Anatolian Neolithic (orange) and CHG/Iran Neolithic (green) in the ADMIXTURE plot. While similar ancestry profiles have been reported for Anatolian and Armenian Chalcolithic and BA individuals, this result suggests the presence of this mixed ancestry north of the Caucasus as early as ~6500 years ago.” ref
Ancient North Eurasian ancestry in Steppe Maykop individuals
“Four individuals from mounds in the grass steppe zone, archaeologically associated with the ‘Steppe Maykop’ cultural complex, lack the Anatolian farmer-related (AF) component when compared to contemporaneous Maykop individuals from the foothills. Instead, they carry a third and fourth ancestry component that is linked deeply to Upper Paleolithic Siberians (maximized in the individual Afontova Gora 3 (AG3) and Native Americans, respectively, and in modern-day North Asians, such as North Siberian Nganasan. To illustrate this affinity with ‘ancient North Eurasians’ (ANE), we also ran PCA with 147 Eurasian and 29 Native American populations. The latter represents a cline from ANE-rich steppe populations such as EHG, Eneolithic individuals, AG3, and Mal’ta 1 (MA1) to modern-day Native Americans at the opposite end. To formally test the excess of alleles shared with ANE/Native Americans, we performed f4-statistics of the form f4(Mbuti, X; Steppe Maykop, Eneolithic steppe), which resulted in significantly positive Z-scores (Z >3) for AG3, MA1, EHG, Clovis, and Kennewick for the ancient populations and many present-day Native American populations.” ref
“Based on these observations, we used qpWave and qpAdm methods to model the number of ancestral sources contributing to the Steppe Maykop individuals and their relative ancestry coefficients. Simple two-way models of Steppe Maykop as an admixture of Eneolithic steppe, AG3, or Kennewick do not fit. However, we could successfully model Steppe Maykop ancestry as being derived from populations related to all three sources (p-value 0.371 for rank 2): Eneolithic steppe (63.5 ± 2.9%), AG3 (29.6 ± 3.4%) and Kennewick (6.9 ± 1.0%). We note that the Kennewick-related signal is most likely driven by the East Eurasian part of Native American ancestry as the f4-statistics (Steppe_Maykop, Fitted Steppe_Maykop; Outgroup1, Outgroup2) show that the Steppe Maykop individuals share more alleles not only with Karitiana but also with Han Chinese.” ref
Characterizing the Caucasus ancestry profile
“The Maykop period, represented by 12 individuals from eight Maykop sites (Maykop, n = 2; a cultural variant ‘Novosvobodnaya’ from the site Klady, n = 4; and Late Maykop, n = 6) in the northern foothills appears homogeneous. These individuals closely resemble the preceding Eneolithic Caucasus individuals and present a continuation of the local genetic profile. This ancestry persisted in the following centuries at least until ~3100 years ago (1100 cal BCE), as revealed by individuals from Kura-Araxes from both the northeast (Velikent, Dagestan) and the South Caucasus (Kaps, Armenia), as well as MBA/LBA individuals (e.g. Kudachurt, Marchenkova Gora) from the north. Overall, this Caucasus ancestry profile falls among the ‘Armenian and Iranian Chalcolithic’ individuals and is indistinguishable from other Kura-Araxes individuals (Armenian EBA) on the PCA plot (Fig. 2), suggesting a dual origin involving Anatolian/Levantine and Iran Neolithic/CHG ancestry, with only minimal EHG/WHG contribution possibly as part of the AF ancestry.” ref
Admixture f3-statistics of the form f3(X, Y; target) with the Caucasus cluster as target resulted in significantly negative Z scores (Z < −3) when CHG (or AG3 in Late Maykop) were used as one and Anatolian farmers as the second potential source. We also used qpWave to determine the number of streams of ancestry and found that a minimum of two is sufficient. We then tested whether each temporal/cultural group of the Caucasus cluster could be modelled as a simple two-way admixture by exploring all possible pairs of sources in qpWave. We found support for CHG as one source and AF ancestry or a derived form such as is found in southeastern Europe as the other. We focused on mixture models of proximal sources (Fig. 4b) such as CHG and Anatolian Chalcolithic for all six groups of the Caucasus cluster (Eneolithic Caucasus, Maykop and Late Makyop, Maykop-Novosvobodnaya, Kura-Araxes, and Dolmen LBA), with admixture proportions on a genetic cline of 40–72% Anatolian Chalcolithic related and 28–60% CHG related.” ref
“When we explored Romania_EN and Bulgaria_Neolithic individuals as alternative southeast European sources (30–46% and 32–49%), the CHG proportions increased to 54–70% and 51–68%, respectively. We hypothesize that alternative models, replacing the Anatolian Chalcolithic individual with yet unsampled populations from eastern Anatolia, South Caucasus or northern Mesopotamia, will likely also provide a fit to some of the tested Caucasus groups. Models with Iran Neolithic as substitute for CHG could also explain the data in a two-way admixture with the combination of Armenia Chalcolithic or Anatolia Chalcolithic as the other source. However, models replacing CHG with EHG received no support, indicating no strong influence for admixture from the adjacent steppe to the north. We also found no direct evidence of EHG or WHG ancestry in Caucasus groups, but observed that Kura-Araxes and Maykop-Novosvobodnaya individuals had likely received additional Iran Chalcolithic-related ancestry (24.9% and 37.4%, respectively).” ref
Characterising the Steppe ancestry profile
“Individuals from the North Caucasian steppe associated with the Yamnaya cultural formation (5300–4400 years ago, 3300–2400 calBCE) appear genetically almost identical to previously reported Yamnaya individuals from Kalmykia immediately to the north, the middle Volga region, Ukraine, and to other BA individuals from the Eurasian steppes who share the characteristic ‘steppe ancestry’ profile as a mixture of EHG and CHG-related ancestry. These individuals form a tight cluster in PCA space (Fig. 2) and can be shown formally to be a mixture by significantly negative admixture f3-statistics of the form f3(EHG, CHG; target). This cluster also involves individuals of the North Caucasus culture (4800–4500 years ago, 2800–2500 calBCE) in the piedmont steppe, who share the steppe ancestry profile, as do individuals from the Catacomb culture in the Kuban, Caspian and piedmont steppes (4600–4200 years ago, 2600–2200 calBCE), which succeeded the Yamnaya horizon.” ref
“The individuals of the MBA post-Catacomb horizon (4200–3700 years ago, 2200–1700 calBCE) such as Late North Caucasus and Lola cultures represent both ancestry profiles common in the North Caucasus: individuals from the mountain site Kabardinka show a typical steppe ancestry profile, whereas individuals from the site Kudachurt 90 km to the west or our most recent individual from the western LBA Dolmen culture (3400–3200 BP, 1400–1200 calBCE) retain the ‘southern’ Caucasus profile. In contrast, one Lola culture individual resembles the ancestry profile of the Steppe Maykop individuals.” ref
Admixture into the steppe zone from the south
“Evidence for interaction between the Caucasus and the Steppe clusters is visible in our genetic data from individuals associated with the later Steppe Maykop phase around 5300–5100 years ago. These ‘outlier’ individuals were buried in the same mounds as those with steppe and in particular Steppe Maykop ancestry profiles but share a higher proportion of AF ancestry visible in the ADMIXTURE plot and are also shifted towards the Caucasus cluster in PC space (Fig. 2d). This observation is confirmed by formal D-statistics. By modelling Steppe Maykop outliers successfully as a two-way mixture of Steppe Maykop and representatives of the Caucasus cluster, we can show that these individuals received additional ‘Anatolian and Iranian Neolithic ancestry’, most likely from contemporaneous sources in the south. We used ALDER to estimate an average admixture time for the observed farmer-related ancestry in Steppe Maykop outliers of 20 generations or 560 years ago.” ref
Anatolian farmer-related ancestry in steppe groups
“Eneolithic Samara individuals form a cline in PC space running from EHG to CHG (Fig. 2d), which is continued by the newly reported Eneolithic steppe individuals. However, the trajectory of this cline changes in the subsequent centuries. Here we observe a cline from Eneolithic_steppe towards the Caucasus cluster. We can qualitatively explain this ‘tilting cline’ by developments south of the Caucasus, where Iranian and AF ancestries continue to mix, resulting in a blend that is also observed in the Caucasus cluster, from where it could have spread onto the steppe. The first appearance of ‘combined farmer-related ancestry’ in the steppe zone is evident in Steppe Maykop outliers. However, PCA results suggest that Yamnaya and later groups of the West Eurasian steppe carry also some farmer-related ancestry as they are slightly shifted towards ‘European Neolithic groups’ in PC2 (Fig. 2d) compared to the preceding Eneolithic steppe individuals. The ‘tilting cline’ is also confirmed by admixture f3-statistics, which provide statistically significant negative values for AG3 and any AF group as the two sources. Using f– and D-statistics we also observe an increase in farmer-related ancestry (both Anatolian and Iranian) in our Steppe cluster, distinguishing the Eneolithic steppe from later groups. In addition, we find the Caucasus cluster or Levant/AF groups to share more alleles with Steppe groups than with EHG or Samara_Eneolithic. MLBA groups such as Poltavka, Andronovo, Srubnaya, and Sintashta show a further increase of AF ancestry consistent with previous studies, reflecting different processes not directly related to events in the Caucasus.” ref
“Reserchers then used qpWave and qpAdm to explore the number of ancestry sources for the AF component to evaluate whether geographically proximate groups contributed plausibly to the subtle shift of Eneolithic ancestry in the steppe towards Neolithic groups. Specifically, we tested whether any of the Eurasian steppe ancestry groups can be successfully modelled as a two-way admixture between Eneolithic steppe and a population X derived from Anatolian- or Iranian farmer-related ancestry, respectively. Surprisingly, we found that a minimum of four streams of ancestry is needed to explain all eight steppe ancestry groups tested (Fig. 2). Importantly, our results show a subtle contribution of both AF ancestry and WHG-related ancestry (Fig. 4), likely brought in through MN/LN farming groups from adjacent regions in the West. A direct source of AF ancestry can be ruled out. At present, due to the limits of our resolution, we cannot identify a single best source population. However, geographically proximal and contemporaneous groups such as Globular Amphora and Eneolithic groups from the Black Sea area (Ukraine and Bulgaria), representing all four distal sources (CHG, EHG, WHG, and Anatolian_Neolithic), are among the best-supported candidates. Applying the same method to the subsequent North Caucasian Steppe groups such as Catacomb, (Late) North Caucasus confirms this pattern.” ref
“Using qpAdm with Globular Amphora as a proximate surrogate population, we estimated the contribution of AF ancestry into Yamnaya and other steppe groups. We find that Yamnaya Samara individuals have 13.2 ± 2.7% and Ukraine or Caucasus Yamnaya individuals 16.6 ± 2.9% AF ancestry—statistically indistinguishable proportions. Substituting Globular Amphora with Iberia Chalcolithic does not alter the results profoundly. This suggests that the source population was a mixture of AF ancestry and a minimum of 20% WHG ancestry, a genetic profile shared by many European MN/LN and Chalcolithic individuals of the 3rd millennium BCE analyzed thus far. To account for potentially un-modelled ancestry from the Caucasus groups, we added ‘Eneolithic Caucasus’ as an additional source to build a three-way model. We found that Yamnaya Caucasus, Yamnaya Ukraine Ozera, North Caucasus and Late North Caucasus had likely received additional ancestry (6–40%) from nearby Caucasus groups. This suggests a more complex and dynamic picture of steppe ancestry groups through time, including the formation of a local variant of steppe ancestry in the North Caucasian steppe from the local Eneolithic, a contribution of Steppe Maykop groups, and population continuity between the early Yamnaya period and the MBA (5300–3200 years ago, 3300–2200 calBCE).” ref
Insights from micro-transects through time
“The availability of multiple individuals from one burial mounds allowed us to test genetic continuity on a micro-transect level. By focusing on two kurgans (Marinskaya 5 and Sharakhalsun 6) with four and five individuals, respectively, we observe that the genetic ancestry varied through time, alternating between the Steppe and Caucasus ancestries, suggesting a shifting genetic border between the two genetic clusters. We also detected various degrees of kinship between individuals buried in the same mound, which supports the view that particular mounds reflected genealogical lineages. Overall, we observe a balanced sex ratio within our sites across the individuals tested (Supplementary Note 4).” ref
A joint model of ancient populations of the Caucasus region
“The fitted qpGraph model recapitulates the genetic separation between the Caucasus and Steppe groups with the Eneolithic steppe individuals deriving more than 60% of ancestry from EHG and the remainder from a CHG-related basal lineage, whereas the Maykop group received about 86.4% from CHG, 9.6% Anatolian farming related ancestry, and 4% from EHG. The Yamnaya individuals from the Caucasus derived the majority of their ancestry from Eneolithic steppe individuals, but also received about 16% from Globular Amphora-related farmers (Fig. 5).” ref

“Admixture Graph modelling of the population history of the Caucasus region. We started with a skeleton tree without admixture, including Mbuti, Loschbour and MA1. We grafted onto this EHG, CHG, Globular_Amphora, Eneolithic_steppe, Maykop, and Yamnaya_Caucasus, adding them consecutively to all possible edges in the tree and retaining only graph solutions that provided no differences of |Z| > 3 between fitted and estimated statistics. The lowest Z-score for this graph is |Z| = 2.824. We note that the maximum discrepancy is f4(MA1, Maykop; EHG, Eneolithic_steppe) = −3.369 if we do not add the 4% EHG ancestry to Maykop. Drifts along edges are multiplied by 1000 and dashed lines represent admixture, Full size image.” ref
“Our data from the Caucasus region cover a 3000-year interval of prehistory, during which we observe a genetic separation between the groups in the northern foothills and those groups of the bordering steppe regions in the north (i.e. the ‘real’ steppe). We have summarised these broadly as Caucasus and Steppe groups in correspondence with eco-geographic vegetation zones that characterise the socio-economic basis of the associated archaeological cultures. When compared to present-day human populations from the Caucasus, which show a clear separation into North and South Caucasus groups along the Great Caucasus mountain range (Fig. 2d), our new data highlight a different situation during the BA. The fact that individuals buried in kurgans in the North Caucasian piedmont zone are more closely related to ancient individuals from regions further south in today’s Armenia, Georgia and Iran results in two main observations.” ref
“First, sometime after the BA present-day North Caucasian populations must have received additional gene-flow from steppe populations that now separates them from southern Caucasians, who largely retained the BA ancestry profile. The archaeological and historic records suggest numerous incursions during the subsequent Iron Age and Medieval times, but ancient DNA from these time periods will be needed to test this directly. Second, our results reveal that the Caucasus was no barrier to human movement in prehistory. Instead the interface of the steppe and northern mountain ecozones could be seen as a transfer zone of cultural innovations from the south and the adjacent Eurasian steppes to the north (Supplementary Note 1). The latter is best exemplified by the two Steppe Maykop outlier individuals, which carry additional AF ancestry, for which the contemporaneous piedmont Maykop individuals present likely candidates for the source of this ancestry. This might also explain the regular presence of ‘Maykop-style artefacts’ in burials that share Steppe Eneolithic traditions and are genetically assigned to the Steppe group. Hence the diverse ‘Steppe Maykop’ group indeed represents the mutual entanglement of Steppe and Caucasus groups and their cultural affiliations in this interaction sphere.” ref
“Concerning the influences from the south, our oldest dates from the immediate Maykop predecessors Darkveti-Meshoko (Eneolithic Caucasus) indicate that the Caucasus genetic profile was present north of the range ~6500 years ago, 4500 calBCE. This is in accordance with the Neolithization of the Caucasus, which had started in the flood plains of South Caucasian rivers in the 6th millennium BCE, from where it spread across to the West/Northwest during the following millennium. It remains unclear whether the local CHG ancestry profile (Kotias Klde and Satsurblia in today’s Georgia) was also present in the North Caucasus region before the Neolithic. However, if we take the CHG ancestry as a local baseline and the oldest Eneolithic Caucasus individuals from our transect as a proxy for the local Late Neolithic ancestry, we notice a substantial increase in AF ancestry. This in all likelihood reflects the process of Neolithization, which also brought this type of ancestry to Europe. As a consequence, it is possible that Neolithic groups could have reached the northern foothills earlier. Hence, additional sampling from older individuals would be desirable to fill this temporal and spatial gap.” ref
We show that the North Caucasus piedmont region was genetically connected to the south at the time of the eponymous grave mound of Maykop. Even without direct ancient DNA data from northern Mesopotamia, our results suggest an increased assimilation of Chalcolithic individuals from Iran, Anatolia, and Armenia and those of the Eneolithic Caucasus during 6000–4000 calBCE, and thus likely also intensified cultural connections. It is possible that the cultural and genetic basis of Maykop were formed within this sphere of interaction (Fig. 4). In fact, the Maykop phenomenon was long understood as the terminus of expanding Mesopotamian civilisations. It has been further suggested that along with these influences the key technological innovations in western Asia that had revolutionised the late 4th millennium BCE had ultimately also spread to Europe. An earlier connection in the late 5th millennium BCE, however, allows speculations about an alternative archaeological scenario: was the cultural exchange mutual and did e.g. metal rich areas such as the Caucasus contribute substantially to the development and transfer of these innovations?” ref
“Within the 3000-year interval covered in this study, we observe a degree of genetic continuity within each cluster, albeit occasionally interspersed by subtle gene-flow between the two clusters as well as from outside sources. Moreover, our data show that the northern flanks were consistently linked to the Near East and had received multiple streams of gene flow from the south during the Maykop, Kura-Araxes, and late phase of the North Caucasus culture. Interestingly, this renewed appearance of the southern genetic make-up in the foothills corresponds to a period of climatic deterioration (known as 4,200 years ago event) in the steppe zone, that put a halt to the exploitation of the steppe zone for several hundred years. Further insight arises from individuals that were buried in the same kurgan but in different time periods, as highlighted in the two kurgans Marinskaya 5 and Sharakhalsun 6. Here, we recognize that the distinction between Steppe and Caucasus (Fig. 1) is not strict but rather reflects a shifting border of genetic ancestry through time, possibly due to climatic/vegetation shifts and/or cultural factors linked to subsistence strategies or social exchange. Thus, the occurrence of Steppe ancestry in the northern foothills likely coincides with the range expansion of Yamnaya pastoralists. However, more time-stamped data from this region will be needed to provide details on the dynamics of this contact zone.” ref
“An important observation is that Eneolithic Samara and Eneolithic steppe individuals directly north of the Caucasus had initially not received AF gene flow. Instead, the Eneolithic steppe ancestry profile shows an even mixture of EHG- and CHG ancestry, suggesting an effective cultural and genetic border between the contemporaneous Eneolithic populations, notably Steppe and Caucasus. Due to the temporal limitations of our dataset, we currently cannot determine whether this ancestry is stemming from an existing natural genetic gradient running from EHG far to the north to CHG/Iran in the south or whether this is the result of Iranian/CHG-related ancestry reaching the steppe zone independently and prior to a stream of AF ancestry, where they mixed with local hunter-gatherers that carried only EHG ancestry.” ref
“All later steppe groups, starting with Yamnaya, deviate from the EHG-CHG admixture cline towards European populations in the West. We show that these individuals had received AF ancestry, in line with published evidence from Yamnaya individuals from Ukraine (Ozera) and Bulgaria. In the North Caucasus, this genetic contribution could have occurred through immediate contact with Caucasus groups or further south. An alternative source, explaining the increase in WHG-related ancestry, would be contact with contemporaneous Chalcolithic/EBA farming groups at the western periphery of the Yamnaya distribution area, such as Globular Amphora and Cucuteni–Trypillia from Ukraine, which have been shown to carry AF ancestry.” ref
“Archaeological arguments are consonant with both scenarios. Contact between early Yamnaya and late Maykop groups is suggested by Maykop impulses seen in early Yamnaya complexes. A western sphere of interaction is evident from striking resemblances of imagery inside burial chambers of Central Europe and the Caucasus, and similarities in geometric decoration patterns in stone cist graves in the Northern Pontic steppe, on stone stelae in the Caucasus, and on pottery of the Eastern Globular Amphora Culture, which links the eastern fringe of the Carpathians and the Baltic Sea. This overlap of symbols implies a late 4th millennium BCE communication and interaction network that operated across the Black Sea area involving the Caucasus, and later also early Globular Amphora groups in the Carpathians and east/central Europe. The role of early Yamnaya groups within this network is still unclear. However, this interaction zone predates any direct influence of Yamnaya groups in Europe, or the succeeding formation of the Corded Ware, and its persistence opens the possibility of subtle gene-flow from farmers at the eastern border of arable lands into the steppe, several centuries before the massive range expansions of pastoralist groups that reached Central Europe in the mid-3rd millennium BCE.” ref
“A surprising discovery was that Steppe Maykop individuals from the eastern desert steppes harboured a distinctive ancestry component that relates them to Upper Palaeolithic Siberians (AG3, MA1) and Native Americans. This is exemplified by the more commonly East Asian features such as the derived EDAR allele, which has also been observed in HG from Karelia and Scandinavia. The additional affinity to East Asians suggests that this ancestry is not derived directly from ANE but from a yet-to-be-identified ancestral population in north-central Eurasia with a wide distribution between the Caucasus, the Ural Mountains, and the Pacific coast, of which we have discovered the so far southwestern-most and also youngest genetic representatives.” ref
“The insight that the Caucasus mountains served as a corridor for the spread of CHG ancestry north but also for subtle later gene-flow from the south allows speculations on the postulated homelands of Proto-Indo-European (PIE) languages and documented gene-flows that could have carried a consecutive spread of both across West Eurasia. This also opens up the possibility of a homeland of PIE south of the Caucasus, and could offer a parsimonious explanation for an early branching off of Anatolian languages, as shown on many PIE tree topologies. Geographically conceivable are also Armenian and Greek, for which genetic data support an eastern influence from Anatolia or the southern Caucasus, and an Indo-Iranian offshoot to the east. However, latest ancient DNA results from South Asia suggest an LMBA spread via the steppe belt. Irrespective of the early branching pattern, the spread of some or all of the PIE branches would have been possible via the North Pontic/Caucasus region and from there, along with pastoralist expansions, to the heart of Europe. This scenario finds support from the well attested and widely documented ‘steppe ancestry’ in European populations and the postulate of increasingly patrilinear societies in the wake of these expansions.” ref
Streams of ancestry and inference of mixture proportions
The researchers used qpWave and qpAdm as implemented in ADMIXTOOLS with the option ‘allsnps: YES’ to test whether a set of test populations is consistent with being related via N streams of ancestry from a set of outgroup populations and estimate mixture proportions for a Test population as a combination of N ‘reference’ populations by exploiting (but not explicitly modeling) shared genetic drift with a set of outgroup populations: Mbuti.DG, Ust_Ishim.DG, Kostenki14, MA1, Han.DG, Papuan.DG, Onge.DG, Villabruna, Vestonice16, ElMiron, Ethiopia_4500BP.SG, Karitiana.DG, Natufian, Iran_Ganj_Dareh_Neolithic. The “DG” samples are extracted from high coverage genomes sequenced as part of the Simons Genome Diversity Project. For some analyses, we used an extended set of outgroup populations, including some of the following additional ancient populations to constrain standard errors: WHG, EHG, and Levant Neolithic.” ref
Jews and Arabs Share Genetic Link to Ancient Canaanites, Study Finds
“Today’s Jews and Arabs in Israel, Jordan, Lebanon, and parts of Syria get half their ancestry from Bronze Age Levantines, who descended from a mix of locals and migrants from Iran or the Caucasus. Most of today’s Jewish and Arabic-speaking populations share a strong genetic link to the ancient Canaanites, a study by an international team of archaeologists and geneticists has found. These modern-day groups in Israel, Jordan, Lebanon, and parts of Syria share a large part of their ancestry, in most cases more than half, with the people who lived in the Levant during the Bronze Age more than 3,000 years ago, the study concludes.” ref
Kura–Araxes culture: Genetic Link to Ancient Canaanites
“The Kura–Araxes culture (also named Kur–Araz culture, Mtkvari–Araxes culture, Early Transcaucasian culture, Shengavitian culture) was an archaeological culture that existed from about 4000 BCE (or 6,000 years ago) until about 2000 BCE (or 4,000 years ago), which has traditionally been regarded as the date of its end; in some locations it may have disappeared as early as 2600 or 2700 BCE. The earliest evidence for this culture is found on the Ararat plain; it spread north in the Caucasus by 3000 BCE. Altogether, the early Transcaucasian culture enveloped a vast area approximately 1,000 km by 500 km, and mostly encompassed the modern-day territories of the South Caucasus (except western Georgia), northwestern Iran, the northeastern Caucasus, eastern Turkey, and as far as Syria.” ref
“The name of the culture is derived from the Kura and Araxes river valleys. Some local variations of the Kura–Araxes culture are sometimes known as Shengavitian, Karaz (Erzurum), Pulur (after a site renamed later as Sakyol), and Yanik Tepe (Iranian Azerbaijan, near Lake Urmia) cultures. It gave rise to the Khirbet Kerak-ware culture found in the Levant and Trialeti culture of the South Caucasus and Armenian Highlands. In Nakhchivan region and nearby areas, Kura-Araxes culture was followed by Nakhchivan culture.” ref
“This civilization was characterized by an agricultural sedentary economy with more than a thousand settlements covering the fertile riverside valleys, high plateaus and high mountain zones of the Armenian highlands and neighboring regions. The Early Bronze Age artificial hill-settlements were characterized by multiple cultural layers, which in some places spread to tens of meters (Mokhrablur of Nakhijevan, Norsun-Tepe). The formative processes of the Kura-Araxes cultural complex, and the date and circumstances of its rise, have been long debated.” ref
“Shulaveri-Shomu culture preceded the Kura–Araxes culture in the area. There were many differences between these two cultures, so the connection was not clear. Later, it was suggested that the Sioni culture of eastern Georgia possibly represented a transition from the Shulaveri to the Kura-Arax cultural complex. At many sites, the Sioni culture layers can be seen as intermediary between Shulaver-Shomu-Tepe layers and the Kura-Araxes layers. This kind of stratigraphy warrants a chronological place of the Sioni culture at around 4000 BCE.” ref
“Some scholars consider the Kartli and Kakheti areas as key to forming the earliest phase of the Kura–Araxes culture. To a large extent, this appears as an indigenous culture of Caucasus that was formed over a long period, and at the same time incorporating foreign influences. There are some indications (such as at Arslantepe) of the overlapping in time of the Kura-Araxes and Uruk cultures; such contacts may go back even to the Middle Uruk period. Some scholars have suggested that the earliest manifestation of the Kura-Araxes phenomenon should be dated at least to the last quarter of the 5th millennium BCE. This is based on the recent data from Ovçular Tepesi, a Late Chalcolithic settlement located in Nakhchivan by the Arpaçay river.” ref
Kura-Araxes Expansion
Kura-Araxes Iranian sites
“There are many important Kura-Araxes sites located along the Araxes river, south of which is modern Iran. Kultepe, Azerbaijan, in Nakhchivan Autonomous Republic close to Araxes river, has already been known for a long time. In Iran, the excavations especially accelerated in the 21st century. Kul Tepe Jolfa, a related site, is seen as an important gateway for Kura-Araxes culture on its way south towards Lake Urmia area. Ancient obsidian trade and distribution are particularly revealing of cultural connections, and Kul Tepe Jolfa was an important transshipment point for the movement of Caucasian obsidian to Iranian sites. This trade was probably conducted by groups of mobile pastoralists from Iran. Also in Iran downstream the Araxes river, two additional sites from the same period have recently been excavated, Kohne Pasgah and Kohne Tepesi. Many other Iranian sites have come to light recently and have been excavated already.” ref
“The economy was based on farming and livestock-raising (especially of cattle and sheep). They grew grain and orchard crops, and are known to have used implements to make flour. They raised cattle, sheep, goats, dogs, and in later phases, horses. Before the Kura-Araxes period, horse bones were not found in Transcaucasia. Later, beginning about 3300 BCE, they became widespread, with signs of domestication. There is evidence of trade with Mesopotamia as well as Asia Minor. It is, however, considered above all to be indigenous to the Caucasus, and its major variants characterized (according to Caucasus historian Amjad Jaimoukha) later major cultures in the region.” ref
“It was at this period that the irrigation systems built on the slopes of the Aragats and Geghama mountains were formed, and at the sources of canals, artificial water pools and springs, dragon stones (vishapakar) made from one piece basalt were erected. Another prerequisite for unprecedented economic development was copper production. A large number of weapon and tools made of arsenical bronze are attested by both the finds near Yerevan and the stone and clay molds found in various ancient sites of Shengavit civilization (Shengavit, Margahovit, etc.). In the Shengavit ancient site, weight standards similar to those used in the Levant were found, which testify the Armenia’s involvement in the newly formed international trade relations in the Early Bronze Age. These weapons that are found were arrows, daggers, battle axes, spears and other weapons made of obsidian, flint, bone and bronze.” ref
“Archaeological evidence of inhabitants of the Kura–Araxes culture showed that ancient settlements were found along the Hrazdan river, as shown by drawings at a mountainous area in a cave nearby. Structures in settlements have not revealed much differentiation, nor was there much difference in size or character between settlements, facts that suggest they probably had a poorly developed social hierarchy for a significant stretch of their history. Some, but not all, settlements were surrounded by stone walls. Among the settlements with an area of 1-10 ha, the central ones were surrounded by fortified walls built of stone (Shengavit, Garni, Persi, Khorenia-Javakhk) and of mud-brick (Mokhrablur, Goy-tepe, Gudaberteke), with artificial puddles (Norabats, Kvatskhelebi, Khizannat-gora). The Shengavit fortified wall is noteworthy with its stone foundations, reinforced with rectangular masonry walls and a tiled secret passage leading to Hrazdan river. The central urban areas, which are characterized by dense construction (Shengavit, Mokhrablur, etc.), were surrounded by satellite residences.” ref
“The Kura-Araxes built mud-brick houses, originally round, but later developing into subrectangular designs with structures of just one or two rooms, multiple rooms centered around an open space, or rectilinear designs. The mud-brick was made from sandy clay with the help of mold forms and dried in the sun. It was the main building material from which 10 to 11 walls, temples, residential and economic buildings, and hydro-engineering structures were built. The foundations of the houses were made of river stones, cracked or unprocessed basalt (Shengavit, Harich, Karaz, Amiranis-gora, etc.), on which mud-brick walls were raised. Round buildings with a diameter of 4-10 m and rectangular floorplans were widespread. The latter had a flat, log roof, and the buildings with round floorplan had primitive roofs of “hazarashen” type, covered with clay mud-bricks with reed, with a skylights hole in the centre of the roof, that solved the issues of light and ventilation of the house (Shengavit, Mokhrablur). The floors were of rammed earth. There were also plaster-covered, up to 10 cm thick (Shengavit) and red-painted (Garakepek-Tepe) floors. There are found samples of attempts to enliven the monotonous clay walls with decoration and with the shaped arrangement of bricks of different colors and to enlighten the monotonous appearance of the walls with various colors (Shengavit, Mokhrablur in Nakhijevan, Yanik-tepe, etc.).” ref
“At some point, the culture’s settlements and burial grounds expanded out of lowland river valleys and into highland areas. Although some scholars have suggested that this expansion demonstrates a switch from agriculture to pastoralism and that it serves as possible proof of a large-scale arrival of Indo-Europeans, facts such as that settlement in the lowlands remained more or less continuous suggest merely that the people of this culture were diversifying their economy to encompass crop and livestock agriculture. Shengavit Settlement is a prominent Kura-Araxes site in present-day Yerevan area in Armenia. It was inhabited from approximately 3,200 to 2,500 BCE cal. Later on, in the Middle Bronze Age, it was used irregularly until 2200 BCE cal. The town occupied an area of six hectares, which is large for Kura-Araxes sites.” ref
“In the earliest phase of the Kura–Araxes culture, metal was scarce. In comparison, the preceding Leilatepe culture‘s metalwork tradition was far more sophisticated. Especially after 3000 BCE, a significant increase in the use of metal objects occurred at Kura-Araxes sites. Also the variation in copper alloys increased during this time. The rich tomb of a woman at Kvazchela is a good example of this, which is quite similar to the ‘royal tomb’ from Arslantepe. The use of an arsenical component up to 25% in copper objects resulted in a shiny greyish, silvery colour. So it’s quite possible that these unusually high arsenical alloys were intended to imitate silver. The Kura–Araxes culture would later display “a precocious metallurgical development, which strongly influenced surrounding regions”. They worked copper, arsenic, silver, gold, tin, and bronze. Their metal goods were widely distributed, from the Volga, Dnieper and Don–Donets river systems in the north to Syria and Palestine in the south and Anatolia in the west.” ref
“Their pottery was distinctive. The spread of their pottery along trade routes into surrounding cultures was much more impressive than any of their achievements domestically. It was painted black and red, using geometric designs. Examples have been found as far south as Syria and Israel, and as far north as Dagestan and Chechnya. The spread of this pottery, along with archaeological evidence of invasions, suggests that the Kura-Araxes people may have spread outward from their original homes and, most certainly, had extensive trade contacts. Jaimoukha believes that its southern expanse is attributable primarily to Mitanni and the Hurrians.” ref
“According to Giulio Palumbi (2008), the typical red-black ware of Kura–Araxes culture originated in eastern Anatolia, and then moved on to the Caucasus area. But then these cultural influences came back to Anatolia mixed in with other cultural elements from the Caucasus. Köhne Shahar is a very large Kura-Araxes archaeological site in Iran located close to the Turkish border. Situated in a narrow valley at the altitude of 1905m asl, it was excavated in 2012-2014. It was located on the trade route between Iran and Anatolia.” ref
“Viticulture and wine-making were widely practised in the area from the earliest times. Viticulture even goes back to the earlier Shulaveri-Shomu culture. The earliest evidence of domesticated grapes in the world has been found at Gadachrili Gora, near the village of Imiri, Marneuli Municipality, in southeastern Republic of Georgia; carbon-dating points to the date of about 6000 BCE. Grape pips dating back to the V-IVth millennia BCE were found in Shulaveri; others dating back to the IVth millennium BCE were found in Khizanaant Gora—all in this same ‘Shulaveri area’ of the Republic of Georgia. A theory has been suggested by Stephen Batiuk that the Kura-Araxes folk may have spread Vitis vinifera vine and wine technology to the “Fertile Crescent“—to Mesopotamia and the Eastern Mediterranean. The spread of the wine-goblet form, such as represented by the Khirbet Kerak ware, is clearly associated with these peoples. The same applies to the large ceramic vessels used for grape fermentation.” ref
Kura-Araxes Religion
“The bearers of the Shengavit culture had a complex religious system. In the central part of the Mokhrablur settlement III construction horizon, a volume-spatial creation was uncovered: a structure-tower with a rectangular plan (7,4×5,5 m) constructed with hard tuff, in the eastern part of which a 3,9 m long one-piece basalt altar was placed. Near this stone structure, clay buildings and ash pits were uncovered, in which the ashes of the sacred hearths had been accumulated. Many tufa idols and clay hearths were excavated in the Shengavit settlement.” ref
“In 2012 year, a complex cult system was excavated-a room with a rectangular plan, designed specifically for ritual ceremonies, inside which a clay altar decorated with relief ornaments on its front was uncovered. A statue of an idol was affixed into the altar and goblets for libation were placed in front of the heart. To the right from the stairs, leading to the semi-subterranean room of the shrine, two clay–packed basins were found, in which the ashes from sacred fires were kept. A phallic pendant-idol was found in the shrine, which was the identifying symbol of the priestess. The adjacent room of the complex reflects household activities of the time. A similar cultic complex was found at the ancient site of Pulur (Sakyol). At the inside of the religious structures, terracotta cult hearths that were unique to the Shengavit culture were located in the altars front. They had diameters of up to one meter, with the edges of the inner space resembling a ship bow divided into three parts, with the upper platforms were red-painted and decorated with geometric figures. Statuettes of women and men and worshiped animals like horses, bulls and rams were found near these hearths. The horseshoe-shaped mobile shrines with ram protomes, threelegged pedestals, phallus-shaped pendant figures were also of religious nature.” ref
Kura-Araxes Burial customs
“One of the spheres of religious practice was the burial ritual. Outside the settlements, burial grounds were formed in their immediate vicinity. Both individual tombs with earthen and tiled walls, as well as wide ancestral tombs with stone walls were revealed, in which the deceased (Joghaz) of the upper-class family were sequentially buried. In the final stage of this civilization, collective burials were performed, which included human sacrifices. These tombs contain numerous artifacts that indications of social stratification: gold and silver jewelry, bronze tools and weapons, imported valuable items. Inhumation practices are mixed. Flat graves are found but so are substantial kurgan burials, the latter of which may be surrounded by dolmen. This points to a heterogeneous ethno-linguistic population (see section below).” ref
“Analyzing the situation in the Kura-Araxes period, T. A. Akhundov notes the lack of unity in funerary monuments, which he considers more than strange in the framework of a single culture; for the funeral rites reflect the deep culture-forming foundations and are weakly influenced by external customs. There are non-kurgan and kurgan burials, burials in-ground pits, in stone boxes and crypts, in the underlying ground strata, and on top of them, using both the round and rectangular burials; there are also substantial differences in the typical corpse position. Burial complexes of Kura–Araxes culture sometimes also include cremation.” ref
“Here one can come to the conclusion that the Kura–Araxes culture developed gradually through a synthesis of several cultural traditions, including the ancient cultures of the Caucasus and nearby territories. Late Kura-Araxes sites often featured Kurgans of greatly varying sizes, with larger, wealthier kurgans surrounded by smaller kurgans containing less wealth. These kurgans also contained a wide assortment metalworks. This trend suggests the eventual emergence of a marked social hierarchy. Their practice of storing relatively great wealth in burial kurgans was probably a cultural influence from the more ancient civilizations of the Fertile Crescent to the south.” ref
“In the 3rd millennium BCE, one particular group of mounds of the Kura–Araxes culture is remarkable for their wealth. This was the final stage of culture’s development. These burial mounds are known as the Martqopi (or Martkopi) period mounds. Those on the left bank of the river Alazani are often 20–25 meters high and 200–300 meters in diameter. They contain especially rich artifacts, such as gold and silver jewelry. While it is unknown what languages were present in Kura-Araxes, the two most widespread theories suggest a connection with Hurro-Urartian and/or Anatolian languages. The Kartvelian and Northeast Caucasian languages were likely spoken in the region as well. A 2023 study employing Bayesian linguistic phylogenetics in conjunction with archaeological, ethnoecological, and human population genetic data suggests that Proto-Karto-Zan (i.e. Proto-Georgian-Zan) prior to its split into Georgian and Zan was spoken by pre-Kura-Araxes and Kura-Araxes farmers that thrived in the watershed of Mtkvari (Kura) River during the Copper and Bronze Ages.” ref
Leyla-Tepe culture
“The Leyla-Tepe culture (Azerbaijani: Leylatəpə mədəniyyəti) of the South Caucasus belongs to the Chalcolithic era. It got its name from the site in the Agdam District of modern-day Azerbaijan. Its settlements were distributed on the southern slopes of Central Caucasus, from 3800 until 3200 BCE. Monuments of the Leyla-Tepe were first located in the 1980s by I. G. Narimanov, a Soviet archaeologist. In the 2000s, attention to the monuments was inspired by the risk of their damage due to the construction of the Baku–Tbilisi–Ceyhan pipeline and the South Caucasus pipeline.” ref
“The settlement of Leyla-Tepe is located on the Karabakh plain at the northwestern outskirts of the small village of Eyvazlı, Agdam, close to the town of Quzanlı in the Aghdam District of Azerbaijan. It was discovered in 1984 by I. Narimanov. It is at an altitude of 100 m above sea level. The semi-dry bed of the Khachinchay River is located 8-10 km to the north. The settlement was originally found as a round hill, 50-60 m in diameter and 2 m in height. Its surface was damaged by modern agriculture, but fragments of ancient ceramics were found on the surface. The thickness of the cultural deposits on the site is about 2 m. Research was carried out at the site, but it was suspended because of the military operations in this region.” ref
“The monument is a single period site without visible cultural changes. Four construction horizons had been identified, of which the upper one was almost completely destroyed by plowing; the lower level was not yet excavated. Few ash pits or refuse accumulations have been found. All buildings examined at the Leilatepe settlement are rectangular in plan. They were erected without a foundation, on the levelled ground surface. The walls were made of rectangular mud bricks laid evenly in horizontal rows with the use of mortar. In the same area of the Aghdam District in the Karabakh valley, there are also some other sites belonging to Leyla-Tepe culture. They are Chinar-Tepe, Shomulu-Tepe (near Mirəşelli village), and Abdal-Aziz-Tepe. Aghdam District had been disputed in recent Nagorno-Karabakh wars.” ref
“The Leyla-Tepe culture is also attested at Böyük Kəsik in the lower layers of this settlement. The inhabitants apparently buried their dead in ceramic vessels. Similar amphora burials in the South Caucasus are found in the Western Georgian Jar-burial culture, that is mostly of a much later date. Jar burials are attested in many parts of the world as early as 4500 BC. The ancient Poylu II settlement was discovered in the Aghstafa District of modern day Azerbaijan during the construction of the Baku–Tbilisi–Ceyhan pipeline. The lowermost layer dates to the early fourth millennium BC, attesting a multilayer settlement of Leyla-Tepe culture.” ref
“Among the sites associated with this culture, the Soyugbulag kurgans or barrows are of special importance. The excavation of these kurgans, located in Kaspi Municipality, in central Georgia, demonstrated an unexpectedly early date of such structures on the territory of Azerbaijan. They were dated to the beginning of the 4th millennium BC. The culture has also been linked to the north Ubaid period monuments, in particular, with the settlements in the Eastern Anatolia Region (Arslantepe, Coruchu-tepe, Tepechik, etc.). The settlement is of a typical Western-Asian variety, with the dwellings packed closely together and made of mud bricks with smoke outlets.” ref
“It has been suggested that the Leyla-Tepe were the founders of the Maykop culture. According to Catherine Marro (2022), Maykop/Majkop culture “is certainly connected with the Leyla Tepe complex”. An expedition to Syria by the Russian Academy of Sciences revealed the similarity of the Maykop and Leyla-Tepe artifacts with those found in 1988–2000 while excavating the ancient city of Tel Khazneh I, dating from the 4th millennium BCE. Leyla-Tepe pottery is very similar to the ‘Chaff-Faced Ware’ of the northern Syria and Mesopotamia. It is especially well attested at Amuq F phase. Similar pottery is also found at Kultepe, Azerbaijan.” ref
“In 2012, the important site of Galayeri, belonging to the Leyla-Tepe archaeological culture, was investigated by Najaf Museibli. It is located in the Qabala District of modern day Azerbaijan. The location is in the area of the Qabala International Airport, about 20km south from the city of Qabala (Gabala). Structures consisting of clay layers are typical; no mud-brick walls have been detected at Galayeri. Almost all findings have Eastern Anatolian Chalcolithic characteristics. The closest analogues of the Galayeri clay constructions are found at Arslantepe/Melid VII in Temple C. The site was further investigated in 2018.” ref
“The archaeological material of the Galayeri settlement is very similar to the finds at the settlements of Böyük Kəsik I, and Poylu II, as well as the Leyla Tepe site. Especially the ceramics are similar at all these sites. Small metal tools finds indicate the production of metal here 6 thousand years ago. Also the remains of a very early potter’s wheel have been found. Radiocarbon dating indicates that Galayeri settlement goes back to the beginning of the 4th millennium BC, which is also supported by the archaeological artifacts found at the settlement.” ref
“The development of Leyla-Tepe culture in the Caucasus marked the early appearance of extractive copper metallurgy. According to research published in 2017, this development that occurred in the second half of the 5th millennium BC preceded the appearance of metallurgy in Mesopotamia. In recent past, the development of copper metallurgy in the Caucasus was attributed to migrants from Uruk arriving around 4500 BCE, or perhaps rather to the pre-Uruk traditions, such as the late Ubaid period, and Ubaid-Uruk phases. Leyla-Tepe metalwork tradition was seen as very sophisticated right from the beginning, and it featured many bronze items. Later, the quality of metallurgy declined somewhat with the advent of the Kura–Araxes culture. Only during the latter stages of Kura–Araxes culture, there was again an improvement in quality.” ref

ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref
Earth diver mythology or something similar??? Could be. In a way, snails are a kind of mound shape, thus similar to turtle shells, both may represent a mound of creation in the earth-diver myth. In Peru, there were snail shells, and snail shells are also used in the earth diver.
My thoughts on Dolmen origins and migrations, as well as Snail Shell Middens or Snail Burials/Turtle Shell Burials, and links from “Y-DNA R (R1a, R1b, and R2a)” migrations, maybe R2a leading to Proto-Indo-European, transferring it to R1b, taking it to the steppe 7,500 years ago.
Religion is a cultural product. So, it has been part of the human experience, similar to languages, from before we left Africa, spreading humanity across the world.


ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref
“The earth-diver is a common character in various traditional creation myths. In these stories, a supreme being usually sends an animal (most often a type of bird, but also crustaceans, insects, and fish in some narratives) into the primal waters to find bits of sand or mud with which to build habitable land.” ref
Axis Mundi Mythology– cosmic axis, world axis, world pillar, center of the world, mound/mountain of creation, or “World/Cosmic tree,” or “Eagle and Serpent tree.” ref, ref
“The World Turtle, also called the Cosmic Turtle or the World-bearing Turtle, is a mytheme of a giant turtle (or tortoise) supporting or containing the world. It occurs in Hindu mythology, Chinese mythology, and the mythologies of some of the indigenous peoples of the Americas.” ref

Yekaterinovsky Cape, a link between the Samara culture and early Khvalynsk
“We already had conflicting information about the elite individual from the Yekaterinovsky Cape and the materials of his grave, which seemed quite old:
For the burial of 45 in the laboratory of the University of Pennsylvania, a 14C date was obtained: PSUAMS-2880 (Sample ID 16068)> 30 kDa gelatin Russia. 12, Ekaterinovka Grave 45 14C age (years ago) 6,325 ± 25 δ 13C (‰) –23.6 δ15 N (‰) 14.5. The results of dating suggest chronological proximity with typologically close materials from Yasinovatsky and Nikolsky burial grounds. The date obtained also precedes the existing dates for the Khvalynsk culture, which, given the dominance of Mariupol traits of the burial rite and inventory, confirms its validity. However, the date obtained for human bones does not exclude the possibility of a “reservoir effect” when the age can increase three or more centuries.” ref
Now the same date is being confirmed by the latest study published on the site, by Korolev, Kochkina, and Stachenkov (2019) and it seems it is really going to be old. Abstract (in part the official one, in part newly translated for clarity):
For the first time, pottery of the Early Eneolithic burial ground Ekaterinovsky Cape is published. Ceramics were predominantly located on the sacrificial sites in the form of compact clusters of fragments. As a rule, such clusters were located above the burials, sometimes over the burials, some were sprinkled with ocher. The authors have identified more than 70 vessels, some of which have been partially reconstructed. Ceramic was made with inclusion of the crushed shell into molding mass. The rims of vessels had the thickened «collar»; the bottoms had a rounded shape. The ornament was located on the rims and the upper part of the potteries. Fully decorated vessels are rare. The vessels are ornamented with prints of comb and rope stamps, with small pits. A particularity of ceramics ornamentation is presented by the imprints of soft stamps (leather?) or traces of leather form for the making of vessels. The ornamentation, made up of «walking comb» and incised lines, was used rarely as well as the belts of pits made decoration under «collar» of a rim.Some features of the ceramics decoration under study relate it with ceramics of the Khvalynsk culture. The ceramics of Ekaterinovsky Cape burial ground is attributed by the authors to the Samara culture. The ceramic complex under study has proximity to the ceramics from Syezzhe burial ground and the ceramics of the second phase of Samara culture. The chronological position is determined by the authors as a later period than the ceramics from the Syezzhe burial ground, and earlier than the chronological position of ceramics of the Ivanovka stage of the Samara culture and the Khvalynsk culture.” ref
More specifically:
Based on ceramic fragments from a large vessel from a cluster of sq.m. 14, the date received was: SPb-2251–5673 ± 120 BP. The second date was obtained in fragments from the aggregation [see picture above] from the cluster of sq.m. 45–46: SPb-2252–6372 ± 100 BP. The difference in dating indicates that the process of determining the chronology of the burial ground is far from complete, although we note that the earlier date almost coincided with the date obtained from the human bone from individual 45 (Korolev, Kochkina, Stashenkov, 2018, p. 300).” ref
“Therefore, the ceramics of the burial ground Ekaterinovsky Cape possess an originality that determines the chronological position of the burial ground between the earliest materials of the burial type in Syezzhe and the Khvalynsk culture. Techno-typological features of dishes make it possible to attribute it to the Samara culture at the stage preceding the appearance of Ivanovska-Khvalynsk ceramics.” ref
“It seems that this site showed cultural influences from the upstream region near the Kama-Vyatka interfluve, too, according to Korolev, Kochkina, Stashenkov, and Khokhlov (2018):
In 2017, excavation of burial ground Ekaterinovsky Cape were continued, located in the area of the confl uence of the Bezenchuk River in the Volga River. During the new excavations, 14 burials were studied. The skeleton of the buried were in a position elongated on the back, less often – crooked on the back with knees bent at the knees. In one burial (No. 90), a special position of the skeleton was recorded. In the burial number 90 in the anatomical order, parts of the male skeleton. This gave grounds for the reconstruction of his original position in a semi-sitting position with the support of elbows on the bottom of the pit. Noteworthy inventory: on the pelvic bones on the left lay a bone spoon, near the right humerus, the pommel of a cruciform club was found.A conclusion is made about the high social status of the buried. The results of the analysis of the burial allow us to outline the closest circle of analogies in the materials of Khvalynsky I and Murzikhinsky burial grounds.” ref
“To sum up, it seems that the relative dates we have used until now have to be corrected: older Khvalynsk I Khvalynsk II individuals, supposedly dated ca. 5200-4000 BCE (most likely after 4700 BCE), and younger Yekaterinovsky individuals, supposedly of the fourth quarter of the 5th millennium (ca. 4250-4000 BCE), are possibly to be considered, in fact, roughly reversed, if not chronologically, at least culturally speaking. Interestingly, this gives a new perspective to the presence of a rare fish- or reptile-headed pommel-scepter, which would be natural in a variable period of expansion of the horse and horse-related symbolism, a cultural trait rooted in the Samara culture attested in Syezzhe before the unification of the symbol of power under the ubiquitous Khvalynsk-Suvorovo horse-headed scepters and related materials.” ref
The Khvalynsk chieftain
“If the reported lineages from Yekaterinovsky Cape are within the R1b-P297 tree, but without further clades, as Yleaf comparisons may suggest, there is not much change to what we have, and R1b-M269 could actually represent a part of the local population, but also incomers from the south (e.g. the north Caspian steppe hunter-gatherers like Kairshak), the east (with hunter-gatherer pottery), or the west near the Don River (in contact with Mariupol-related cultures, as the authors inferred initially from material culture). Just like R1a-M417 became incorporated into the Sredni Stog groups after the Novodanilovka-Suvorovo expansion, probably as incoming hunter-gatherer pottery groups from the north admixing with peoples of “Steppe ancestry”, R1b-M269 lineages might have expanded explosively only during the Repin expansion, and maybe (like R1b-L51 later) they formed just a tiny part of the clans that dominated the steppe during the Khvalynsk-Novodanilovka community.” ref
“On the other hand, the potential finding of various R1b-M269/L23 samples in Yekaterinovsky Cape (including an elite individual) would suggest now, as it was supported in the original report by Mathieson et al. (2015), that these ancient R1b lineages found in the Volga – Ural region are in fact most likely all R1b-M269 without enough coverage to obtain proper SNP calls, which would simplify the picture of Neolithic expansions (yet again). From the supplementary materials:
10122 / SVP35 (grave 12). Male (confirmed genetically), age 20-30, positioned on his back with raised knees, with 293 copper artifacts, mostly beads, amounting to 80% of the copper objects in the combined cemeteries of Khvalynsk I and II. Probably a high-status individual, his Y-chromosome haplotype, R1b1, also characterized the high-status individuals buried under kurgans in later Yamnaya graves in this region, so he could be regarded as a founder of an elite group of patrilineally related families. His MtDNA haplotype H2a1 is unique in the Samara series.” ref
“This remarkable Khvalynsk chieftain, whose rich assemblage may correspond to the period of domination of the culture all over the Pontic-Caspian steppes, has been consistently reported as of hg. R1b-L754 in all publications, including Wang et al. (2018/2019) tentative SNP calls in the supplementary materials (obtained with Yleaf, as the infamous Narasimhan et al. 2018 samples), but has been variously reported by amateurs as within the R1b-M73, R1b-V88, or (lately) R1b-V1636 trees, which makes it unlikely that quality of the sample is allowing for a proper SNP call. The fact that Mathieson et al. (2015) considered it a member of the R1b-M269 clans appearing later in Yamna seems on point right now, especially if samples from Yekaterinovka are all within this tree. The relevance of R1b-L23 in the expansion of Repin and Yamna is reminiscent of the influence of successful clans among Yamna offshoots, such as Bell Beakers, and among Bell Beaker offshoots during the Bronze Age all over Europe.” ref
“Taking these younger expansions as example, it seems quite likely based on cultural links that (at least part of) the main clans of Khvalynsk were of R1b-M269 lineage, stemming from a R1b-dominated Samara culture, in line with the known succeeding expansions and the expected strictly patriarcal and patrilineal society of Proto-Indo-Europeans, which would have exacerbated the usual reduction in Y-chromosome haplogroup variability that happens during population expansions, and the aversion towards foreign groups while the culture lasted.” ref
“The finding of R1b-L23 in Yekaterinovka, associated with the Samara culture, before or during the Khvalynsk expansion, and close to the Khvalynsk site, would make this Khvalynsk chieftain most likely a member of the M269 tree (paradoxically, the only R1b-L754 branch amateurs have not yet reported for it). Similarly, the sample of a “Samara hunter-gatherer” of Lebyazhinka, of hg. R1b-P297, could also be under this tree, just like most R1b-M269 from Yamna are downstream from R1b-L23, and most reported R1b-M269 or R1b-L23 from Bell Beakers are under R1b-L151.” ref
“On the other hand, we know of the shortcomings of attributing a haplogroup expansion to the best known rulers, such as the famous lineages previously wrongly attributed to Niall of the Nine Hostages or Genghis Khan. The known presence of R1b-V1636 up to modern Greeks would be in line with an ancient steppe expansion that we know will show up during the Neolithic, although it could also be a sign of a more recent migration from the Caucasus. The presence of a sister clade of R1b-L23, R1b-PF7562, among modern Balkan populations, may also be attributed to a pre-Yamna steppe expansion.” ref
“I reckon that even informal reports on SNP calls, like any other analyses, should be offered in full: not only with a personal or automatic estimation of the result, but with a detailed explanation of the good, dubious, and bad calls, alternatives to that SNP estimation, and a motivated reasoning of why one branch should be preferred over others. Downloading a sample and giving an instruction using a free software tool is never enough, as it became crystal clear recently for the hilariously biased and flawed qpAdm reports on Dutch Bell Beakers as the ‘missing link’ between Corded Ware and Bell Beakers…” ref
“Another example I can recall is the report of a R1a-Z93 subclade in the R1a-M417 sample ca. 4000 BC from Alexandria, which seems rather unlikely, seeing how this subclade must have split and expanded explosively with R1a-Z645 to the east with eastern Corded Ware groups, i.e. 1,000 years later, just like Z282 lineages expanded mainly to the north-east. But then again, as with the Khvalynsk chieftain, I have only seen indirect reports of that supposed SNP (including Y26+!), so we should just stick with its officially reported R1a-M417 lineage. This upstream haplogroup was, in fact, repeated with Yleaf’s tentative estimates in Wang et al. (2019) supplementary materials…” ref
“The combination of inexperienced, biased, or simply careless design, analyses, and reports, including SNP calls and qpAdm analyses (whether in forums or publications), however well-intentioned (or not) they might be, are hindering a proper analysis of data, adding to the difficulties we already have due to the scarcity of samples, their limited coverage, and the lack of proper context. Some people like to repeat ad nauseam that archaeology and/or linguistics are ‘not science’ whenever they don’t fit their beliefs and myths based on haplogroup and/or ancestry. But it’s becoming harder and harder to rely on certain genetic data, too, and on their infinite changing interpretations, much more than it is to rely on linguistic and archaeological research, including data, assessments, and discussions that are open for anyone to review…if one is truly interested in them.” ref

“The Paleolithic dog was a Late Pleistocene canine. They were directly associated with human hunting camps in Europe over 30,000 years ago and it is proposed that these were domesticated. They are further proposed to be either a proto-dog and the ancestor of the domestic dog or an extinct, morphologically and genetically divergent wolf population. There are a number of recently discovered specimens which are proposed as being Paleolithic dogs, however, their taxonomy is debated. These have been found in either Europe or Siberia and date 40,000–17,000 years ago. They include Hohle Fels in Germany, Goyet Caves in Belgium, Predmosti in the Czech Republic, and four sites in Russia: Razboinichya Cave in the Altai Republic, Kostyonki-8, Ulakhan Sular in the Sakha Republic, and Eliseevichi 1 on the Russian plain.” ref
1. 40,000–35,500 years ago Hohle Fels, Schelklingen, Germany
2. 36,500 years ago Goyet Caves, Samson River Valley, Belgium
3. 33,500 years ago Razboinichya Cave, Altai Mountains, (Russia/Siberia)
4. 33,500–26,500 years ago Kostyonki-Borshchyovo archaeological complex, (Kostenki site) Voronezh, Russia
5. 31,000 years ago Predmostí, Moravia, Czech Republic
6. 26,000 years ago Chauvet Cave, Vallon-Pont-d’Arc, Ardèche region, France
7. 17,300–14,100 years ago Dyuktai Cave, northern Yakutia, Siberia
8. 17,000–16,000 years ago Eliseevichi-I site, Bryansk Region, Russian Plain, Russia
9. 16,900 years ago Afontova Gora-1, Yenisei River, southern Siberia
10. 14,223 years ago Bonn–Oberkassel, Germany
11. 13,500 years ago Mezine, Chernigov region, Ukraine
12. 13,000 years ago Palegawra, (Zarzian culture) Iraq
13. 12,800 years ago Ushki I, Kamchatka, eastern Siberia
14. 12,790 years ago Nanzhuangtou, China
15. 12,300 years ago Ust’-Khaita site, Baikal region, Siberia
16. 12,000 years ago Ain Mallaha (Eynan) and HaYonim terrace, Israel
17. 10,150 years ago Lawyer’s Cave, Alaska, USA
18. 9,000 years ago Jiahu site, China
19. 8,000 years ago Svaerdborg site, Denmark
20. 7,425 years ago Lake Baikal region, Siberia
21. 7,000 years ago Tianluoshan archaeological site, Zhejiang province, China ref
1. 40,000–35,500 years ago Hohle Fels, Schelklingen, Germany
“Canid maxillary fragment. The size of the molars matches those of a wolf, the morphology matches a dog. Proposed as a Paleolithic dog. The figurine Venus of Hohle Fels was discovered in this cave and dated to this time.” ref
2. 36,500 years ago Goyet Caves, Samson River Valley, Belgium
“The “Goyet dog” is proposed as being a Paleolithic dog. The Goyet skull is very similar in shape to that of the Eliseevichi-I dog skulls (16,900 years ago) and to the Epigravettian Mezin 5490 and Mezhirich dog skulls (13,500 years ago), which are about 18,000 years younger. The dog-like skull was found in a side gallery of the cave, and Palaeolithic artifacts in this system of caves date from the Mousterian, Aurignacian, Gravettian, and Magdalenian, which indicates recurrent occupations of the cave from the Pleniglacial until the Late Glacial. The Goyet dog left no descendants, and its genetic classification is inconclusive because its mitochondrial DNA (mDNA) does not match any living wolf nor dog. It may represent an aborted domestication event or phenotypically and genetically distinct wolves. A genome-wide study of a 35,000-year-old Pleistocene wolf fossil from northern Siberia indicates that the dog and the modern grey wolf genetically diverged from a common ancestor between 27,000 and 40,000 years ago.” ref
3. 33,500 years ago Razboinichya Cave, Altai Mountains, (Russia/Siberia)
“The “Altai dog” is proposed as being a Paleolithic dog. The specimens discovered were a dog-like skull, mandibles (both sides), and teeth. The morphological classification, and an initial mDNA analysis, found it to be a dog. A later study of its mDNA was inconclusive, with 2 analyses indicating dog and another 2 indicating wolf. In 2017, two prominent evolutionary biologists reviewed the evidence and supported the Altai dog as being a dog from a lineage that is now extinct and that was derived from a population of small wolves that are also now extinct.” ref
4. 33,500–26,500 years ago Kostyonki-Borshchyovo archaeological complex, (Kostenki site) Voronezh, Russia
“One left mandible paired with the right maxilla, proposed as a Paleolithic dog.” ref
5. 31,000 years ago Predmostí, Moravia, Czech Republic
“Three dog-like skulls proposed as being Paleolithic dogs. Predmostí is a Gravettian site. The skulls were found in the human burial zone and identified as Palaeolithic dogs, characterized by – compared to wolves – short skulls, short snouts, wide palates and braincases, and even-sized carnassials. Wolf skulls were also found at the site. One dog had been buried with a bone placed carefully in its mouth. The presence of dogs buried with humans at this Gravettian site corroborates the hypothesis that domestication began long before the Late Glacial. Further analysis of bone collagen and dental microwear on tooth enamel indicates that these canines had a different diet when compared with wolves (refer under diet).” ref
6. 26,000 years ago Chauvet Cave, Vallon-Pont-d’Arc, Ardèche region, France
“50-metre trail of footprints made by a boy of about ten years of age alongside those of a large canid. The size and position of the canid’s shortened middle toe in relation to its pads indicate a dog rather than a wolf. The footprints have been dated by soot deposited from the torch the child was carrying. The cave is famous for its cave paintings. A later study using geometric morphometric analysis to compare modern wolves with modern dog tracks proposes that these are wolf tracks.” ref
7. 17,300–14,100 years ago Dyuktai Cave, northern Yakutia, Siberia
“Large canid remains along with human artifacts. And from a nearby site dating to around 17,200–16,800 Ulakhan Sular, northern Yakutia, Siberia held a fossil dog-like skull similar in size to the “Altai dog”, proposed as a Paleolithic dog.” ref
8. 17,000–16,000 years ago Eliseevichi-I site, Bryansk Region, Russian Plain, Russia
“Two fossil canine skulls proposed as being Paleolithic dogs. In 2002, a study looked at the fossil skulls of two large canids that had been found buried 2 meters and 7 meters from what was once a mammoth-bone hut at this Upper Paleolithic site, and using an accepted morphologically based definition of domestication declared them to be “Ice Age dogs”. The carbon dating gave a calendar-year age estimate that ranged between 16,945 and 13,905 years ago. The Eliseevichi-1 skull is very similar in shape to the Goyet skull (36,000 years ago), the Mezine dog skull (13,500 years ago) and Mezhirich dog skull (13,500 years ago). In 2013, a study looked at the mDNA sequence for one of these skulls and identified it as Canis lupus familiaris i.e. dog. However, in 2015 a study using three-dimensional geometric morphometric analyses indicated the skull is more likely from a wolf. These animals were larger in size than most grey wolves and approached the size of a Great Dane.” ref
9. 16,900 years ago Afontova Gora-1, Yenisei River, southern Siberia
“Fossil dog-like tibia, proposed as a Paleolithic dog. The site is on the western bank of the Yenisei River about 2,500 km southwest of Ulakhan Sular, and shares a similar timeframe to that canid. A skull from this site described as dog-like has been lost in the past, but there exists a written description of it possessing a wide snout and a clear stop, with a skull length of 23 cm that falls outside of those of wolves.” ref
10. 14,223 years ago Bonn–Oberkassel, Germany
“The “Bonn-Oberkassel dog“. Undisputed dog skeleton buried with a man and woman. All three skeletal remains were found sprayed with red hematite powder. The consensus is that a dog was buried along with two humans. Analysis of mDNA indicates that this dog was a direct ancestor of modern dogs. Domestic dog.” ref
11. 13,500 years ago Mezine, Chernigov region, Ukraine
“Ancient dog-like skull proposed as being a Paleolithic dog. Additionally, ancient wolf specimens were found at the site. Dated to the Epigravettian period (17,000–10,000 years ago). The Mezine skull is very similar in shape to the Goyet skull (36,000 years ago), the Eliseevichi-1 dog skulls (16,900 years ago), and the Mezhirich dog skull (13,500 years ago). The Epigravettian Mezine site is well known for its round mammoth bone dwelling. Taxonomy uncertain.” ref
12. 13,000 years ago Palegawra, (Zarzian culture) Iraq
“The fossil jaw and teeth of a domesticated dog, recovered from a cave in Iraq, have been found to be about 14,000 years old. The bone was found in a shallow cave with a number of stone tools suggesting that its keepers were hunters. The scientists who found and studied the bone speculated that the animal served either as a hunting dog in the field or as a watchdog back at the cave or perhaps as both.” ref, ref
13. 12,800 years ago Ushki I, Kamchatka, eastern Siberia
“Complete skeleton buried in a buried dwelling. Located 1,800 km to the southeast from Ulakhan Sular. Domestic dog.” ref
14. 12,790 years ago Nanzhuangtou, China
“31 fragments including a complete dog mandible.” ref
15. 12,300 years ago Ust’-Khaita site, Baikal region, Siberia
“Sub-adult skull located 2,400 km southwest of Ulakhan Sular and proposed as a Paleolithic dog. Also a somewhat close find at 12,450 years old mummified dog carcass. The “Black Dog of Tumat” was found frozen into the ice core of an oxbow lake steep ravine at the middle course of the Syalaah River in the Ust-Yana region. DNA analysis confirmed it as an early dog.” ref The Archaeology of Ushki Lake, Kamchatka, and the Pleistocene Peopling of the Americas: The Ushki Paleolithic sites of Kamchatka, Russia, have long been thought to contain information critical to the peopling of the Americas, especially the origins of Clovis. New radiocarbon dates indicate that human occupation of Ushki began only 13,000 calendar years ago-nearly 4000 years later than previously thought. Although biface industries were widespread across Beringia contemporaneous to the time of Clovis in western North America, these data suggest that late-glacial Siberians did not spread into Beringia until the end of the Pleistocene, perhaps too recently to have been ancestral to proposed pre-Clovis populations in the Americas.” ref
16. 12,000 years ago Ain Mallaha (Eynan) and HaYonim terrace, Israel
“Three canid finds. A diminutive carnassial and a mandible, and a wolf or dog puppy skeleton buried with a human during the Natufian culture. These Natufian dogs did not exhibit tooth-crowding. The Natufian culture occupied the Levant, and had earlier interred a fox together with a human in the Uyun al-Hammam burial site, Jordan dated 17,700–14,750 years ago.” ref
17. 10,150 years ago Lawyer’s Cave, Alaska, USA
“Bone of a dog, oldest find in North America. DNA indicates a split from Siberian relatives 16,500 years ago, indicating that dogs may have been in Beringia earlier. Lawyer’s Cave is on the Alaskan mainland east of Wrangell Island in the Alexander Archipelago of southeast Alaska.” ref
18. 9,000 years ago Jiahu site, China
“Eleven dog interments. Jaihu is a Neolithic site 22 kilometers north of Wuyang in Henan Province.” ref Most archaeologists consider the Jaihu site to be one of the earliest examples of the Peiligang culture. Settled around 7000 BCE or around 9,000 years ago, the site was later flooded and abandoned around 5700 BCE or around 7,700 years ago. At one time, it was “a complex, highly organized Chinese Neolithic society”, home to at least 250 people and perhaps as many as 800. The important discoveries of the Jiahu archaeological site include the Jiahu symbols, possibly an early example of proto-writing, carved into tortoise shells and bones; the thirty-three Jiahu flutes carved from the wing bones of cranes, believed to be among the oldest playable musical instruments in the world; and evidence of alcohol fermented from rice, honey and hawthorn leaves.” ref
19. 8,000 years ago Svaerdborg site, Denmark
“Three different sized dog types recorded at this Maglemosian culture site. Maglemosian (c. 9000 – c. 6000 BCE or around 11,000 to 8,000 years ago) is the name given to a culture of the early Mesolithic period in Northern Europe. In Scandinavia, the culture was succeeded by the Kongemose culture. It appears that they had domesticated the dog. Similar settlements were excavated from England to Poland and from Skåne in Sweden to northern France.” ref, ref
20. 7,425 years ago Lake Baikal region, Siberia
“Dog buried in a human burial ground. Additionally, a human skull was found buried between the legs of a “tundra wolf” dated 8,320 years ago (but it does not match any known wolf DNA). The evidence indicates that as soon as formal cemeteries developed in Baikal, some canids began to receive mortuary treatments that closely paralleled those of humans. One dog was found buried with four red deer canine pendants around its neck dated 5,770 years ago. Many burials of dogs continued in this region with the latest finding at 3,760 years ago, and they were buried lying on their right side and facing towards the east as did their humans. Some were buried with artifacts, e.g., stone blades, birch bark, and antler bone.” ref
21. 7,000 years ago Tianluoshan archaeological site, Zhejiang province, China
“In 2020, an mDNA study of ancient dog fossils from the Yellow River and Yangtze River basins of southern China showed that most of the ancient dogs fell within haplogroup A1b, as do the Australian dingoes and the pre-colonial dogs of the Pacific, but in low frequency in China today. The specimen from the Tianluoshan archaeological site is basal to the entire lineage. The dogs belonging to this haplogroup were once widely distributed in southern China, then dispersed through Southeast Asia into New Guinea and Oceania, but were replaced in China 2,000 years ago by dogs of other lineages.” ref

ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref
An interesting potential dog genetic lineage is connected to a group of ancient canids date to more than 47,000 years ago had separated from the other ancient canids including wolves. Genetic studies of modern dog and wolf populations show origins in East/South Asia and/or the Near East to multiple areas of domestication and/or hybridization with regional wolf breeds.
A 33,000-year-old emerging dog from southern Siberia in the Altai Mountains seems to demonstrate an early domestication. The oldest similar emergence of this behavior seems to be demonstrated a pre-Natufian burial site in Jordan Uyun al-Hammam dated to around 16,500-year-old with elaborate human burials with grave goods as well as include evidence for unique human-animal relationships, seeming to show foxes where used similar to modern dogs demonstrating that the dog like domestication features were not unique to the later Natufians.
Moreover, dog genetics is one way to further demonstrate human migration as well as its oven accompanying religious transfer. While most dogs buried at this time were individual others were placed back-to-back in pairs. Moreover, a general genetic analysis of modern dogs suggests a general origin in southern China approximately 16,000 years ago. The Natufian culture existed in the Levant roughly from 14,500 to 11,500 years. It seems two different human burials at the Ain Mallaha Natufian settlement and Hayonim cave sites include dogs which likely suggest dogs were domesticated by at least by around 12,000 years ago.
In addition, at Ain Mallaha there is a widespread influence of the culture and as always, the presumed religious transfer can be estimated by the presence of obsidian from Turkey and shellfish from the Nile-valley as part of the artifacts found. Furthermore, generally by around 12,000 years ago domestic dogs are presumed to be found from the Levant, Cyprus, Iraq, Northern China, and the Kamchatka peninsula in Far Eastern Russia. A 12,000-year-old tomb in northern Israel held a fifty-year-old woman was buried with a puppy close to her head with her left hand on it seemingly expressing a religious or an emotional connection, possibly some kind of shaman burial. By around 8,000 years ago at Svaerdborg in Denmark there are already three differently sized dog types found.
")
- My Thoughts on Possible Migrations of “R” DNA and Proto-Indo-European?
- Migrations and Changing Europeans Beginning around 8,000 Years Ago
- My “Steppe-Anatolian-Kurgan hypothesis” 8,000/7,000 years ago
- The Dnieper–Donets culture and Asian varieties of Millet from China to the Black Sea region of Europe by 7,022 years ago
- Kurgan 6,000 years ago/dolmens 7,000 years ago: funeral, ritual, and other?
- 7,020 to 6,020-year-old Proto-Indo-European Homeland of Urheimat or proposed home of their Language and Religion


My Speculations are in Comparative Mythologies?
For instance, the mytheme of an ancient belief that is seemingly shared though changed and adapted, a fundamental generic unit of narrative structure seems to be shared a common relation with mountains/ancestors/gods or sacred animals with Sacred Mounds, Mountains, Kurgans, and Pyramids.
Sacred Mounds, Mountains, Kurgans, and Pyramids may hold deep Mythology connections?


ref, ref, ref, ref, ref, ref, ref, ref, ref
“Several linguists and geneticists suggest that the Uralic languages are related to various Siberian languages and possibly also some languages of northern Native Americans. A proposed family is named Uralo-Siberian, it includes Uralic, Yukaghir, Eskimo–Aleut (Inuit), possibly Nivkh, and Chukotko-Kamchatkan. Haplogroup Q is found in nearly all Native Americans and nearly all of the Yeniseian Ket people (90%).” ref, ref
You can find some form of Shamanism, among Uralic, Transeurasian, Dené–Yeniseian, Chukotko-Kamchatkan, and Eskaleut languages.
My speculations of shamanism are its dispersals, after 24,000 to 4,000 years ago, seem to center on Lake Baikal and related areas. To me, the hotspot of Shamanism goes from west of Lake Baikal in the “Altai Mountains” also encompassing “Lake Baikal” and includes the “Amur Region/Watershed” east of Lake Baikal as the main location Shamanism seems to have radiated out from.

“Dené–Caucasian is a discredited language family proposal that includes widely-separated language groups spoken in the Northern Hemisphere: Sino-Tibetan languages, Yeniseian languages, Burushaski and North Caucasian languages in Asia; Na-Dené languages in North America; and the Vasconic languages from Europe (including Basque). A narrower connection specifically between North American Na-Dené and Siberian Yeniseian (the Dené–Yeniseian languages hypothesis) was proposed by Edward Vajda in 2008, and has met with some acceptance within the community of professional linguists. The validity of the rest of the family, however, is viewed as doubtful or rejected by nearly all historical linguists.” ref
“The Dené–Caucasian family tree and approximate divergence dates (estimated by modified glottochronology) proposed by S. A. Starostin and his colleagues from the Tower of Babel project:
- Dené–Caucasian languages [8,700 BCE or around 10,700 years ago]
- Na-Dené languages (Athabascan–Eyak–Tlingit)
- Sino-Vasconic languages [7,900 BCE or around 9,900 years ago]
- Vasconic (see below)
- Sino-Caucasian languages [6,200 BCE or around 8,200 years ago]
- Burushaski
- Caucaso-Sino-Yeniseian [5,900 BCE or around 7,900 years ago]
- North Caucasian languages
- Sino-Yeniseian [5,100 BCE or around 7,100 years ago]
“John D. Bengtson groups Basque, Caucasian and Burushaski together in a Macro-Caucasian (earlier Vasco-Caucasian) family (see the section on Macro-Caucasian below). According to him, it is as yet premature to propose other nodes or subgroupings, but he notes that Sumerian seems to share the same number of isoglosses with the (geographically) western branches as with the eastern ones:
- Dené–Caucasian
- The Macro-Caucasian family
- Basque
- North Caucasian
- Burushaski
- Sumerian
- Sino-Tibetan
- Yeniseian
- Na-Dené” ref
- The Macro-Caucasian family
“It has been conjectured that the North-West Caucasian languages may be genetically related to the Indo-European family, at a time depth of perhaps 12,000 years before the present. This hypothesized proto-language is called Proto-Pontic, but is not widely accepted. There does at least appear to have been extensive contact between the two proto-languages, and the resemblances may be due to this influence. A few linguists have proposed even broader relationships, of which the Dene–Caucasian hypothesis is perhaps the most popular. Dene–Caucasian links the North Caucasian (including Northwest Caucasian), Basque, Burushaski, Yeniseian, Sino-Tibetan, and Na–Dene families. However, this is an even more tentative hypothesis than Nostratic, which attempts to relate Kartvelian, Indo-European, Uralic, and Altaic, etc., and which is widely considered to be undemonstrated.” ref
“Nostratic is a hypothetical language macrofamily including many of the language families of northern Eurasia. Though a historically important proposal, in a contemporary context it is typically considered a fringe theory. Although the exact composition varies based on proponent, it typically comprises Kartvelian, Indo-European and Uralic languages; some languages from the similarly controversial Altaic family; the Afroasiatic languages; as well as the Dravidian languages (sometimes also Elamo-Dravidian).” ref
“Iran Neolithic (Iran_N) individuals dated ~8,500 years ago carried 50% Ancient North Eurasian-derived admixture and 50% Dzudzuana-related admixture, marking them as different from other Near-Eastern and Anatolian Neolithics who didn’t have Ancient North Eurasian admixture. Iran Neolithics were later replaced by Iran Chalcolithics, who were a mixture of Iran Neolithic and Near Eastern Levant Neolithic.” ref
I speculate that possibly this “Iran Neolithic” difference is a later migration relating to Ancient North Eurasian admixture, with the source languages from Siberia (pre-proto-indo-europeain, like some kind of pre/proto-Yeniseian, or Dené–Yeniseian languages/or Dené–Caucasian) that then merged into proto-indo-European languages seen just west of Iran in the Caucasus and East Turkey areas. Also, I speculate that the idea of pottery was likewise brought by these peoples and they, to me could have influenced the creation of the earliest pottery in Tell Hassuna and Jarmo (Iraq).
“Proto-Yeniseian or Proto-Yeniseic is the unattested reconstructed proto-language from which all Yeniseian languages are thought to descend from. It is uncertain whether Proto-Yeniseian had a similar tone/pitch accent system as Ket people, who practiced Shamanism and connected to Tengrism. Many studies about Proto-Yeniseian phonology have been done, however there are still many things unclear about Proto-Yeniseian. The probable location of the Yeniseian homeland is proposed on the basis of geographic names and genetic studies, which suggests a homeland in Southern Siberia.” ref
“Tengri (Old Turkic: 𐰚𐰇𐰚:𐱅𐰭𐰼𐰃, romanized: Kök Teŋri/Teŋiri, lit. ’Blue Heaven’; Old Uyghur: tängri; Middle Turkic: تآنغرِ; Ottoman Turkish: تڭری; Kyrgyz: Теңир; Kazakh: Тәңір; Turkish: Tanrı; Azerbaijani: Tanrı; Bulgarian: Тангра; Proto-Turkic *teŋri / *taŋrɨ; Mongolian script: ᠲᠩᠷᠢ, T’ngri; Mongolian: Тэнгэр, Tenger; Uyghur: تەڭرى tengri ) is the all-encompassing God of Heaven in the traditional Turkic, Yeniseian, Mongolic, and various other nomadic Altaic religious beliefs. Tengri is not considered a deity in the usual sense, but a personification of the universe. However, some qualities associated with Tengri as the judge and source of life, and being eternal and supreme, led European and Muslim writers to identify Tengri as a deity of Turkic and Mongolic peoples. According to Mongolian belief, Tengri’s will (jayayan) may break its own usual laws and intervene by sending a chosen person to earth. It is also one of the terms used for the primary chief deity of the early Turkic and Mongolic peoples. Worship surrounding Tengri is called Tengrism. The core beings in Tengrism are the Sky Father (Tenger Etseg) and the Earth Mother (Umay Ana). It involves ancestor worship, as Tengri was thought to have been the ancestral progenitor of mankind in Turkic regions and Mongolia, shamanism, animism, and totemism.” ref
“Tengrism (also known as Tengriism, Tengerism, or Tengrianism) is a religion originating in the Eurasian steppes, based on shamanism and animism. It generally involves the titular sky god Tengri, who is not considered a deity in the usual sense but a personification of the universe. According to some scholars, adherents of Tengrism view the purpose of life to be in harmony with the universe. It was the prevailing religion of the Göktürks, Xianbei, Bulgars, Xiongnu, Yeniseian, and Mongolic peoples and Huns, as well as the state religion of several medieval states: the First Turkic Khaganate, the Western Turkic Khaganate, the Eastern Turkic Khaganate, Old Great Bulgaria, the First Bulgarian Empire, Volga Bulgaria, Khazaria, and the Mongol Empire. In the Irk Bitig, a ninth century manuscript on divination, Tengri is mentioned as Türük Tängrisi (God of Turks). According to many academics, Tengrism was, and to some extent still is, a predominantly polytheistic religion based on the shamanistic concept of animism, and was first influenced by monotheism during the imperial period, especially by the 12th–13th centuries. Abdulkadir Inan argues that Yakut and Altai shamanism are not entirely equal to the ancient Turkic religion.” ref
“The term also describes several contemporary Turkic and Mongolic native religious movements and teachings. All modern adherents of “political” Tengrism are monotheists. Tengrism has been advocated for in intellectual circles of the Turkic nations of Central Asia (Kyrgyzstan with Kazakhstan) and Russia (Tatarstan, Bashkortostan) since the dissolution of the Soviet Union during the 1990s. Still practiced, it is undergoing an organized revival in Buryatia, Sakha (Yakutia), Khakassia, Tuva and other Turkic nations in Siberia. Altaian Burkhanism and Chuvash Vattisen Yaly are contemporary movements similar to Tengrism. The term tengri (compare with Kami) can refer to the sky deity Tenger Etseg – also Gök Tengri; Sky father, Blue sky – or to other deities. While Tengrism includes the worship of personified gods (tngri) such as Ülgen and Kaira,Tengri is considered an “abstract phenomenon”. In Mongolian folk religion, Genghis Khan is considered one of the embodiments, if not the main embodiment, of Tengri’s will. The forms of the name Tengri (Old Turkic: Täŋri) among the ancient and modern Turkic and Mongolic are Tengeri, Tangara, Tangri, Tanri, Tangre, Tegri, Tingir, Tenkri, Tangra, Teri, Ter, and Ture. The name Tengri (“the Sky”) is derived from Old Turkic: Tenk (“daybreak”) or Tan (“dawn”). Meanwhile, Stefan Georg proposed that the Turkic Tengri ultimately originates as a loanword from Proto-Yeniseian *tɨŋgɨr- “high”. Mongolia is sometimes poetically called the “Land of Eternal Blue Sky” (Mönkh Khökh Tengeriin Oron) by its inhabitants. According to some scholars, the name of the important deity Dangun (also Tangol) (God of the Mountains) of the Korean folk religion is related to the Siberian Tengri (“Heaven”), while the bear is a symbol of the Big Dipper (Ursa Major).” ref
“Tiān (天) is one of the oldest Chinese terms for heaven and a key concept in Chinese mythology, philosophy, and religion. During the Shang dynasty (17th―11th century BCE), the Chinese referred to their highest god as Shàngdì (上帝, “Lord Above”) or Dì (帝, “Lord”). During the following Zhou dynasty, Tiān became synonymous with this figure. Before the 20th century, worship of Tiān was an orthodox state religion of China. In Chinese culture, heaven tends to be “synonymous with order”, “containing the blueprints for creation”, “the mandate by which earthly rulers govern, and the standards by which to measure beauty, goodness, and truth.” Zhou dynasty nobles made the worship of heaven a major part of their political philosophy and viewed it as “many gods” who embodied order and kingship, as well as the mandate of heaven. For the etymology of tiān, Schuessler links it with the Mongolian word tengri “sky, heaven, heavenly deity” or the Tibeto-Burman words taleŋ (Adi) and tǎ-lyaŋ (Lepcha), both meaning “sky”. He also suggests a likely connection between Chinese tiān 天, diān 巔 “summit, mountaintop”, and diān 顛 “summit, top of the head, forehead”, which have cognates such as Zemeic Naga tiŋ “sky”. However, other reconstructions of 天’s OC pronunciation *qʰl’iːn or *l̥ˤi[n] reconstructed a voiceless lateral onset, either a cluster or a single consonant, respectively. Baxter & Sagart pointed to attested dialectal differences in Eastern Han Chinese, the use of 天 as a phonetic component in phono-semantic compound Chinese characters, and the choice of 天 to transcribe foreign syllables, all of which prompted them to conclude that, around 200 CE, 天’s onset had two pronunciations: coronal *tʰ & dorsal *x, both of which likely originated from an earlier voiceless lateral *l̥ˤ.” ref
“In Taoism and Confucianism, Tiān (the celestial aspect of the cosmos, often translated as “Heaven“) is mentioned in relationship to its complementary aspect of Dì (地, often translated as “Earth“). They are thought to maintain the two poles of the Three Realms (三界) of reality, with the middle realm occupied by Humanity (人, rén), and the lower world occupied by demons (魔, mó) and “ghosts”, the damned, (鬼, guǐ). Tiān was variously thought as a “supreme power reigning over lesser gods and human beings” that brought “order and calm…or catastrophe and punishment”, a god, destiny, an “impersonal” natural force that controlled various events, a holy world or afterlife containing other worlds or afterlives, or one or more of these. “Confucianism has a religious side with a deep reverence for Heaven and Earth (Di), whose powers regulate the flow of nature and influence human events.” Yin and yang are also thought to be integral to this relationship and permeate both, as well as humans and man-made constructs. This “cosmos” and its “principles” is something that “[t]he ways of man should conform to, or else” frustration will result. Many Confucianists, both historically and in current times, use the I Ching to divine events through the changes of Tiān and other “natural forces”. Historical and current Confucianists were/are often environmentalists out of their respect for Heaven and the other aspects of nature and the “Principle” that comes from their unity and, more generally, harmony as a whole, which is “the basis for a sincere mind.” The Emperor of China as Tianzi was formerly vital to Confucianism. Mount Tai is seen as a sacred place in Confucianism and was traditionally the most revered place where Chinese emperors offered sacrifices to heaven and earth. Some tiān in Chinese folk religion were thought to be many different or a hierarchy of multiple, sphere-like realms that contained morally ambiguous creatures and spirits such as huli jing and fire-breathing dragons. ” ref
“Paleo-Siberian languages, languages spoken in Asian Russia (Siberia) that belong to four genetically unrelated groups—Yeniseian, Luorawetlan, Yukaghir, and Nivkh.” ref
Proto-Indo-European mythology
“Proto-Indo-European mythology is the body of myths and deities associated with the Proto-Indo-Europeans, speakers of the hypothesized Proto-Indo-European language. Although the mythological motifs are not directly attested – since Proto-Indo-European speakers lived in preliterate societies – scholars of comparative mythology have reconstructed details from inherited similarities found among Indo-European languages, based on the assumption that parts of the Proto-Indo-Europeans’ original belief systems survived in the daughter traditions. The Proto-Indo-European pantheon includes a number of securely reconstructed deities, since they are both cognates – linguistic siblings from a common origin – and associated with similar attributes and body of myths: such as *Dyḗws Ph₂tḗr, the daylight-sky god; his consort *Dʰéǵʰōm, the earth mother; his daughter *H₂éwsōs, the dawn goddess; his sons the Divine Twins; and *Seh₂ul and *Meh₁not, a solar goddess and moon god, respectively. Some deities, like the weather god *Perkʷunos or the herding-god *Péh₂usōn, are only attested in a limited number of traditions – Western (i.e. European) and Graeco-Aryan, respectively – and could therefore represent late additions that did not spread throughout the various Indo-European dialects.” ref
“Some myths are also securely dated to Proto-Indo-European times, since they feature both linguistic and thematic evidence of an inherited motif: a story portraying a mythical figure associated with thunder and slaying a multi-headed serpent to release torrents of water that had previously been pent up; a creation myth involving two brothers, one of whom sacrifices the other in order to create the world; and probably the belief that the Otherworld was guarded by a watchdog and could only be reached by crossing a river. Various schools of thought exist regarding possible interpretations of the reconstructed Proto-Indo-European mythology. The main mythologies used in comparative reconstruction are Indo-Iranian, Baltic, Roman, and Norse, often supported with evidence from the Celtic, Greek, Slavic, Hittite, Armenian, Illyrian, and Albanian traditions as well.” ref
“Early agricultural communities such as Chogha Golan in 10,000 BCE or around 12,000 years ago, along with settlements such as Chogha Bonut (the earliest village in Elam) in 8000 BCE or around 10,000 years ago, began to flourish in and around the Zagros Mountains region in western Iran. Around about the same time, the earliest-known clay vessels and modeled human and animal terracotta figurines were produced at Ganj Dareh, also in western Iran. There are also 10,000-year-old human and animal figurines from Tepe Sarab in Kermanshah Province among many other ancient artifacts.” ref
I also speculate that there may be a connection with this to the earliest pottery in Turkey from Boncuklu Höyük as well.
“12 fired clay samples and an unfired marl sample from the late 9th and early 8th-millennium BCE site of Boncuklu Höyük (8300–7800 cal BCE or around 10,300 to 9,800 years ago) in the Konya Plain, Turkey. The clay vessels from Boncuklu Höyük, an early Neolithic site in central Anatolia, are much earlier than the accepted date for the introduction of pottery in Anatolia, c. 7000 cal BCE or around 9,000 years ago.” ref
Proto-Indo-European (PIE) is the reconstructed common ancestor of the Indo-European language family:
| Subdivisions |
|---|
“Western Iran was inhabited by a population genetically most similar to hunter-gatherers from the Caucasus, but distinct from the Neolithic Anatolian people who later brought food production into Europe. While some degree of cultural diffusion between Anatolia, Western Iran, and other neighboring regions is possible, the genetic dissimilarity between early Anatolian farmers and the inhabitants of Ganj Dareh supports a model in which Neolithic societies in these areas were distinct. The genome of an early Neolithic female from Ganj Dareh, GD13a, from the Central Zagros (Western Iran), dated to 10000-9700 cal years ago, a region located at the eastern edge of the Near East. Ganj Dareh is well known for providing the earliest evidence of herd management of goats beginning at 9,900 years ago. The mitochondrion of GD13a (91.74X) was assigned to haplogroup X, most likely to the subhaplogroup X2, which has been associated with an early expansion from the Near East and has been found in early Neolithic samples from Anatolia, Hungary, and Germany. GD13a did not cluster with any other early Neolithic individual from Eurasia in any of the analyses. Also genetically close to GD13a were ancient samples from Steppe populations (Yamanya & Afanasievo) that were part of one or more Bronze age migrations into Europe, as well as early Bronze age cultures in that continent (Corded Ware), in line with previous relationships observed for the Caucasus Hunter-Gatherers.” ref
“Subclade X2 appears to have undergone extensive population expansion and dispersal around or soon after the Last Glacial Maximum, roughly 20,000 years ago. It is more strongly represented in the Near East, the Caucasus, and southern Europe, and somewhat less strongly present in the rest of Europe. The highest concentrations are found in the Ojibwe (25%), Sioux (15%), Nuu-Chah-Nulth (12%), Georgia (8%), Orkney (7%), and amongst the Druze Assyrian community in Israel (27%). Subclades of X2 are not present in South Americans Amerindian populations. The oldest known human associated with X2 is Kennewick Man, whose c. 9000-year old remains were discovered in Washington State. The lineage of haplogroup X in the Americas is not derived from a European subclade, but rather represents an independent subclade, labeled X2a. The X2a subclade has not been found in Eurasia, and has most likely arisen within the early Paleo-Indian population, at roughly 13,000 years ago. A basal variant of X2a was found in the Kennewick Man fossil (ca. 9,000 years ago). No presence of mt-DNA ancestral to X2a has been found in Europe or the Near East. New World lineages X2a and X2g are not derived from the Old World lineages X2b, X2c, X2d, X2e, and X2f, indicating an early origin of the New World lineages “likely at the very beginning of their expansion and spread from the Near East.” ref
“Although it occurs only at a frequency of about 3% for the total current indigenous population of the Americas, it is a bigger haplogroup in northern North America, where among the Algonquian peoples it comprises up to 25% of mtDNA types. It is also present in lesser percentages to the west and south of this area—among the Sioux (15%), the Nuu-chah-nulth (11%–13%), the Navajo (7%), and the Yakama (5%). In Latin America, Haplotype X6 was present in the Tarahumara 1.8% (1/53) and Huichol 20% (3/15) X6 and X7 was also found in 12% in Yanomani people. Unlike the four main Native American mtDNA haplogroups (A, B, C, D), X is not strongly associated with East Asia. The main occurrence of X in Asia discovered so far is in the Altai people in Siberia. One theory of how the X Haplogroup ended up in North America is that the people carrying it migrated from central Asia along with haplogroups A, B, C, and D, from an ancestor from the Altai Region of Central Asia. Two sequences of haplogroup X2 were sampled further east of Altai among the Evenks of Central Siberia. These two sequences belong to X2* and X2b. It is uncertain if they represent a remnant of the migration of X2 through Siberia or a more recent input.” ref
“Haplogroup X has been found in various other bone specimens that were analysed for ancient DNA, including specimens associated with the Alföld Linear Pottery (X2b-T226C, Garadna-Elkerülő út site 2, 1/1 or 100%), Linearbandkeramik (X2d1, Halberstadt-Sonntagsfeld, 1/22 or ~5%), and Iberia Chalcolithic (X2b, La Chabola de la Hechicera, 1/3 or 33%; X2b, El Sotillo, 1/3 or 33%; X2b, El Mirador Cave, 1/12 or ~8%) cultures. Abel-beth-maachah 2201 was a man who lived between 1014 and 836 BCE during the Levant Iron Age and was found in the region now known as Abel Beth Maacah, Metula, Israel. He was associated with the Galilean cultural group. His direct maternal line belonged to mtDNA haplogroup X2b. Haplogroup X has been found in ancient Assyria and ancient Egyptian mummies excavated at the Abusir el-Meleq archaeological site in Middle Egypt, which date from the late New Kingdom and Roman periods. Fossils excavated at the Late Neolithic site of Kelif el Boroud in Morocco, which have been dated to around 5,000 years old, have also been found to carry the X2 subclade.” ref

Pic ref
Ancient Human Genomes…Present-Day Europeans – Johannes Krause (Video)
Ancient North Eurasian (ANE)
Eastern Hunter-Gatherer (EHG)
Western Hunter-Gatherers (WHG)
Scandinavian Hunter-Gatherer (SHG)
Early European Farmers (EEF)
A quick look at the Genetic history of Europe
“The most significant recent dispersal of modern humans from Africa gave rise to an undifferentiated “non-African” lineage by some 70,000-50,000 years ago. By about 50–40 ka a basal West Eurasian lineage had emerged, as had a separate East Asian lineage. Both basal East and West Eurasians acquired Neanderthal admixture in Europe and Asia. European early modern humans (EEMH) lineages between 40,000-26,000 years ago (Aurignacian) were still part of a large Western Eurasian “meta-population”, related to Central and Western Asian populations. Divergence into genetically distinct sub-populations within Western Eurasia is a result of increased selection pressure and founder effects during the Last Glacial Maximum (LGM, Gravettian). By the end of the LGM, after 20,000 years ago, A Western European lineage, dubbed West European Hunter-Gatherer (WHG) emerges from the Solutrean refugium during the European Mesolithic. These Mesolithic hunter-gatherer cultures are substantially replaced in the Neolithic Revolution by the arrival of Early European Farmers (EEF) lineages derived from Mesolithic populations of West Asia (Anatolia and the Caucasus). In the European Bronze Age, there were again substantial population replacements in parts of Europe by the intrusion of Ancient North Eurasian (ANE) lineages from the Pontic–Caspian steppes. These Bronze Age population replacements are associated with the Beaker culture archaeologically and with the Indo-European expansion linguistically.” ref
“As a result of the population movements during the Mesolithic to Bronze Age, modern European populations are distinguished by differences in WHG, EEF, and ANE ancestry. Admixture rates varied geographically; in the late Neolithic, WHG ancestry in farmers in Hungary was at around 10%, in Germany around 25%, and in Iberia as high as 50%. The contribution of EEF is more significant in Mediterranean Europe, and declines towards northern and northeastern Europe, where WHG ancestry is stronger; the Sardinians are considered to be the closest European group to the population of the EEF. ANE ancestry is found throughout Europe, with a maximum of about 20% found in Baltic people and Finns. Ethnogenesis of the modern ethnic groups of Europe in the historical period is associated with numerous admixture events, primarily those associated with the Roman, Germanic, Norse, Slavic, Berber, Arab and Turkish expansions. Research into the genetic history of Europe became possible in the second half of the 20th century, but did not yield results with a high resolution before the 1990s. In the 1990s, preliminary results became possible, but they remained mostly limited to studies of mitochondrial and Y-chromosomal lineages. Autosomal DNA became more easily accessible in the 2000s, and since the mid-2010s, results of previously unattainable resolution, many of them based on full-genome analysis of ancient DNA, have been published at an accelerated pace.” ref

“The arrival of haplogroup R1a-M417 in Eastern Europe, and the east-west diffusion of pottery through North Eurasia.” https://indo-european.eu/2018/02/the-arrival-of-haplogroup-r1a-m417-in-eastern-europe-and-the-east-west-diffusion-of-pottery-through-north-eurasia/
Ancient North Eurasian https://en.wikipedia.org/wiki/Ancient_North_Eurasian
Ancient North Eurasian/Mal’ta–Buret’ culture haplogroup R* https://en.wikipedia.org/wiki/Mal%27ta%E2%80%93Buret%27_culture
“The arrival of haplogroup R1a-M417 in Eastern Europe, and the east-west diffusion of pottery through North Eurasia.” ref
R-M417 (R1a1a1)
“R1a1a1 (R-M417) is the most widely found subclade, in two variations which are found respectively in Europe (R1a1a1b1 (R-Z282) ([R1a1a1a*] (R-Z282) and Central and South Asia (R1a1a1b2 (R-Z93) ([R1a1a2*] (R-Z93).” ref
R-Z282 (R1a1a1b1a) (Eastern Europe)
“This large subclade appears to encompass most of the R1a1a found in Europe.
- R1a1a1b1a [R1a1a1a*] (R-Z282*) occurs in northern Ukraine, Belarus, and Russia at a frequency of c. 20%.
- R1a1a1b1a3 [R1a1a1a1] (R-Z284) occurs in Northwest Europe and peaks at c. 20% in Norway.
- R1a1a1c (M64.2, M87, M204) is apparently rare: it was found in 1 of 117 males typed in southern Iran.” ref
R1a1a1b2 (R-Z93) (Asia)
“This large subclade appears to encompass most of the R1a1a found in Asia, being related to Indo-European migrations (including Scythians, Indo-Aryan migrations, and so on).
- R-Z93* or R1a1a1b2* (R1a1a2* in Underhill (2014)) is most common (>30%) in the South Siberian Altai region of Russia, cropping up in Kyrgyzstan (6%) and in all Iranian populations (1-8%).
- R-Z2125 occurs at highest frequencies in Kyrgyzstan and in Afghan Pashtuns (>40%). At a frequency of >10%, it is also observed in other Afghan ethnic groups and in some populations in the Caucasus and Iran.
- R-M434 is a subclade of Z2125. It was detected in 14 people (out of 3667 people tested), all in a restricted geographical range from Pakistan to Oman. This likely reflects a recent mutation event in Pakistan.
- R-M560 is very rare and was only observed in four samples: two Burushaski speakers (north Pakistan), one Hazara (Afghanistan), and one Iranian Azerbaijani.
- R-M780 occurs at high frequency in South Asia: India, Pakistan, Afghanistan, and the Himalayas. The group also occurs at >3% in some Iranian populations and is present at >30% in Roma from Croatia and Hungary.” ref
R-M458 (R1a1a1b1a1)
“R-M458 is a mainly Slavic SNP, characterized by its own mutation, and was first called cluster N. Underhill et al. (2009) found it to be present in modern European populations roughly between the Rhine catchment and the Ural Mountains and traced it to “a founder effect that … falls into the early Holocene period, 7.9±2.6 KYA.” M458 was found in one skeleton from a 14th-century grave field in Usedom, Mecklenburg-Vorpommern, Germany. The paper by Underhill et al. (2009) also reports a surprisingly high frequency of M458 in some Northern Caucasian populations (for example 27.5% among Karachays and 23.5% among Balkars, 7.8% among Karanogays and 3.4% among Abazas).” ref
“Migration from Siberia behind the formation of Göbeklitepe: Expert states. People who migrated from Siberia formed the Göbeklitepe, and those in Göbeklitepe migrated in five other ways to spread to the world, said experts about the 12,000-year-old Neolithic archaeological site in the southwestern province of Şanlıurfa.“ The upper paleolithic migrations between Siberia and the Near East is a process that has been confirmed by material culture documents,” he said.” ref
“Semih Güneri, a retired professor from Caucasia and Central Asia Archaeology Research Center of Dokuz Eylül University, and his colleague, Professor Ekaterine Lipnina, presented the Siberia-Göbeklitepe hypothesis they have developed in recent years at the congress held in Istanbul between June 11 and 13. There was a migration that started from Siberia 30,000 years ago and spread to all of Asia and then to Eastern and Northern Europe, Güneri said at the international congress.” ref
“The relationship of Göbeklitepe high culture with the carriers of Siberian microblade stone tool technology is no longer a secret,” he said while emphasizing that the most important branch of the migrations extended to the Near East. “The results of the genetic analyzes of Iraq’s Zagros region confirm the traces of the Siberian/North Asian indigenous people, who arrived at Zagros via the Central Asian mountainous corridor and met with the Göbeklitepe culture via Northern Iraq,” he added.” ref
“Emphasizing that the stone tool technology was transported approximately 7,000 kilometers from east to west, he said, “It is not clear whether this technology is transmitted directly to long distances by people speaking the Turkish language at the earliest, or it travels this long-distance through using way stations.” According to the archaeological documents, it is known that the Siberian people had reached the Zagros region, he said. “There seems to be a relationship between Siberian hunter-gatherers and native Zagros hunter-gatherers,” Güneri said, adding that the results of genetic studies show that Siberian people reached as far as the Zagros.” ref
“There were three waves of migration of Turkish tribes from the Southern Siberia to Europe,” said Osman Karatay, a professor from Ege University. He added that most of the groups in the third wave, which took place between 2600-2400 BCE, assimilated and entered the Germanic tribes and that there was a genetic kinship between their tribes and the Turks. The professor also pointed out that there are indications that there is a technology and tool transfer from Siberia to the Göbeklitepe region and that it is not known whether people came, and if any, whether they were Turkish.” ref
“Around 12,000 years ago, there would be no ‘Turks’ as we know it today. However, there may have been tribes that we could call our ‘common ancestors,’” he added. “Talking about 30,000 years ago, it is impossible to identify and classify nations in today’s terms,” said Murat Öztürk, associate professor from İnönü University. He also said that it is not possible to determine who came to where during the migrations that were accepted to have been made thousands of years ago from Siberia. On the other hand, Mehmet Özdoğan, an academic from Istanbul University, has an idea of where “the people of Göbeklitepe migrated to.” ref
“According to Özdoğan, “the people of Göbeklitepe turned into farmers, and they could not stand the pressure of the overwhelming clergy and started to migrate to five ways.” “Migrations take place primarily in groups. One of the five routes extends to the Caucasus, another from Iran to Central Asia, the Mediterranean coast to Spain, Thrace and [the northwestern province of] Kırklareli to Europe and England, and one route is to Istanbul via [Istanbul’s neighboring province of] Sakarya and stops,” Özdoğan said. In a very short time after the migration of farmers in Göbeklitepe, 300 settlements were established only around northern Greece, Bulgaria, and Thrace. “Those who remained in Göbeklitepe pulled the trigger of Mesopotamian civilization in the following periods, and those who migrated to Mesopotamia started irrigated agriculture before the Sumerians,” he said.” ref







“Rb1-V88 around 8,000 years ago. R1b-V88 coalescence time was estimated between 9,200–5,600 years ago. Researchers suggest R1b-V88 is a paternal genetic record of the proposed mid-Holocene migration of proto-Chadic Afroasiatic speakers through the Central Sahara into the Lake Chad Basin, and geomorphological evidence is consistent with this view.” ref
Sumerians
Genetic footprints of Sumerians: a survey of Y-chromosome and mtDNA variation in the Marsh Arabs of Iraq
“For millennia, the southern part of the Mesopotamia has been a wetland region generated by the Tigris and Euphrates rivers before flowing into the Gulf. This area has been occupied by human communities since ancient times, and the present-day inhabitants, the Marsh Arabs, are considered the population with the strongest link to ancient Sumerians. Analyses of the haplogroups and sub-haplogroups observed in the Marsh Arabs revealed a prevalent autochthonous Middle Eastern component for both male and female gene pools, with weak South-West Asian and African contributions, more evident in mtDNA. A higher male than female homogeneity is characteristic of the Marsh Arab gene pool, likely due to a strong male genetic drift determined by socio-cultural factors (patrilocality, polygamy, unequal male and female migration rates).” ref
“Evidence of genetic stratification ascribable to the Sumerian development was provided by the Y-chromosome data where the J1-Page08 branch reveals a local expansion, almost contemporary with the Sumerian City State period that characterized Southern Mesopotamia. On the other hand, a more ancient background shared with Northern Mesopotamia is revealed by the less represented Y-chromosome lineage J1-M267*. Overall our results indicate that the introduction of water buffalo breeding and rice farming, most likely from the Indian sub-continent, only marginally affected the gene pool of autochthonous people of the region. Furthermore, a prevalent Middle Eastern ancestry of the modern population of the marshes of southern Iraq implies that if the Marsh Arabs are descendants of the ancient Sumerians, also the Sumerians were most likely autochthonous and not of Indian or South Asian ancestry.” ref
“The Near East is well known for its important role in human history, particularly as a theatre for great historical events that changed the face of the world during the Neolithic period. The temperate climate and fertile soil brought by the continuous flooding of the Tigris and Euphrates rivers, made the Mesopotamian region ideal for early revolutions in agriculture and farming. In particular, the southern part of Mesopotamia (the delta between the two rivers in the present day southern Iraq) has been historically known as the Garden of Eden (biblical name) or Sumer Land, the land of Abraham. The Mesopotamian civilization originated around the 4th millennium BCE (around 6,000 years ago) in the low course of the Tigris and Euphrates rivers. This alluvial territory, which emerged progressively by soil sedimentation, attracted different populations from the northern and eastern mountains but, whereas traces of their culture are present in the territory, as documented by the Ubaid-Eridu pottery, nothing is available for their identification.” ref
“Only two groups of populations arrived later and in larger number leaved historical records: Sumerian and Semitic groups. The Sumerians, who spoke an isolated language not correlated to any linguistic family, are the most ancient group living in the region for which we have historical evidence. They occupied the delta between the two rivers in the southern part of the present Iraq, one of the oldest inhabited wetland environments. The Semitic groups were semi-nomadic people who spoke a Semitic language and lived in the northern area of the Syro-Arabian desert breeding small animals. From here, they reached Mesopotamia where they settled among the pre-existing populations. The Semitic people, more numerous in the north, and the Sumerians, more represented in the south, after having adsorbed the pre-existing populations, melted their cultures laying the basis of the western civilization.” ref
“The ancient inhabitants of the marsh areas were Sumerians, who were the first to develop an urban civilization some 5,000 years ago. Although footprints of their great civilization are still evident in prominent archaeological sites lying on the edges of the marshes, such as the ancient Sumerian cities of Lagash, Ur, Uruk, Eridu, and Larsa, the origin of Sumerians is still a matter of debate. With respect to this question, two main scenarios have been proposed: according to the first, the original Sumerians were a group of populations who had migrated from “the Southeast” (India region) and took the seashore route through Arabian Gulf before settling down in the southern marshes of Iraq. The second hypothesis posits that the advancement of the Sumerian civilization was the result of human migrations from the mountainous area of Northeastern Mesopotamia to the southern marshes of Iraq, with ensuing assimilation of the previous populations.” ref
“Over time, the many historical and archaeological expeditions that have been conducted in the marshes have consistently reported numerous parallelisms between the modern and ancient life styles of the marsh people. Details such as home architecture (particular arched reed buildings), food gathering (grazing water buffalos, trapping birds and spearing fish, rice cultivation), and means of transportation (slender bitumen-covered wooden boats, called “Tarada”) are documented as still being practiced by the indigenous population locally named “Ma’dan” or “Marsh Arabs”. This village life-style, which has remained unchanged for seven millennia, suggests a possible link between the present-day marsh inhabitants and ancient Sumerians. However, popular tradition considers the Marsh Arabs as a foreign group, of unknown origin, which arrived in the marshlands when the rearing of water buffalo was introduced to the region.” ref
“Marsh Arabs and Iraqis identified 28 haplogroups, 14 in the marsh sample and 22 in the control Iraqis. Only eight haplogroups were shared by both groups. More than 90% of both Y-chromosome gene pools can be traced back to Western Eurasian components: the Middle Eastern Hg J-M304, the Near Eastern Hgs G-M201, E-M78, and E-M123, while the Eurasian Hgs I-M170 and R-M207 are scarce and less common in the Marsh Arabs than in the control sample. Contributions from eastern Asia, India, and Pakistan, represented by Hgs L-M76, Q-M378, and R2-M124, are detected in the Marsh Arabs, but at a very low frequency.” ref
“Haplogroup J accounts for 55.1% of the Iraqi sample reaching 84.6% in the Marsh Arabs, one of the highest frequencies reported so far. Unlike the Iraqi sample, which displays a roughly equal proportion of J1-M267 (56.4%) and J2-M172 (43.6%), almost all Marsh Arab J chromosomes (96%) belongs to the J1-M267 clade and, in particular, to sub-Hg J1-Page08. Haplogroup E, which characterizes 6.3% of Marsh Arabs and 13.6% of Iraqis, is represented by E-M123 in both groups, and E-M78 mainly in the Iraqis. Haplogroup R1 is present at a significantly lower frequency in the Marsh Arabs than in the Iraqi sample (2.8% vs 19.4%; P < 0.001), and is present only as R1-L23. Conversely the Iraqis are distributed in all the three R1 sub-groups (R1-L23, R1-M17 and R1-M412) found in this survey at frequencies of 9.1%, 8.4% and 1.9%, respectively. Other haplogroups encountered at low frequencies among the Marsh Arabs are Q (2.8%), G (1.4%), L (0.7%), and R2 (1.4%).” ref
“When the PCA was based on mtDNA haplogroup frequencies, Marsh Arabs occupied, together with Iraqi and Saudi Arabian populations, a position in the middle of the plot among three distinct groupings: the first included western Eurasian, the second embraced all the South Asian groups while the third represented the North Africa and South Arabian Peninsula peoples. For both systems, the longitudinal separation operated by the first PC is mainly due to the East-West decreasing frequency of East Asian haplogroups (see for example: Y-chromosome Hgs R2-M124, C-RPS4Y, and H-M69; mtDNA Hgs A, F, D, and G) and the increasing frequencies of the African haplogroups (see for example: Y-chromosome Hgs A-M13, B-M60, E-M35; mtDNA Hgs L1, L2, and L3) while the latitudinal separation operated by the second PC is mainly ascribable to the different distribution of haplogroups most frequent in West Eurasian (Y-chromosome Hgs J-M172, M267 and mtDNA Hgs H and U5), and the African-specific haplogroups (Y-chromosome Hgs A-M13, B-M60, E-M35, and mtDNA Hgs L0-3).” ref
“MtDNA variation: A total of 233 haplotypes and 77 sub-haplogroups have been identified in this survey. Only 26 of the observed sub-haplogroups are shared between the two populations, and most of the remaining are represented by singletons. According to their known or supposed geographic/ethnic origin, in addition to a strong West Eurasian component (77.8% and 84.1% in the Marsh Arabs and Iraqis, respectively), it is possible to recognize contributions from North/East and Sub-Saharan Africa and from East and South Asia.” ref
“West Eurasian mtDNAs observed in this study are approximately equally distributed into macro-Hgs R0, KU, and JT, although with haplogroup and sub-haplogroup differences between the two Iraqi samples. In the Marsh Arabs Hg J prevails (15.2%) followed by Hgs H (12.4%), U (9.7%) and T (7.6%). Conversely, in the control group, the most frequent is Hg H (17.0%), followed by Hgs U (14.8%), T (12.6%), and J (11.9%). Both the less represented N1 and W haplogroups show higher frequencies (marginally significant) in Marsh Arabs. The most frequent macro-Hg R0 includes molecules R0a ((preHV)I), more represented among the Marsh Arabs (6.9% vs 4.0%), HV, observed mainly as HV*, but especially H mtDNAs. Although the majority of the H mtDNAs (7.6% in Marsh Arabs vs 10.8% in Iraqis) did not fall into any of the tested sub-haplogroups, a limited number of H subsets (H1, H5, H6, and H14) have been observed. In particular, while H5 (3.4% vs. 2.8%), H1 (0.7% vs. 1.7%), and H14 (0.7% vs. 1.1%) were found in both groups, H6 was observed only in one subject of the control group.” ref
“Almost all the main U sub-haplogroups and the nested K branch were found in the Iraqi sample, but only a sub-set of them (K1, U3, U4, U5, in addition to the South West Asian U7) were observed in the Marsh Arabs. The nested Hg K, mainly K1, was observed at a comparable frequency in both groups (6.2% in the marshes vs 4.6%). The situation of macro-Hg JT is more complex. Significant differences (P < 0.05) emerged in the distribution of J1 and J2 sub-clades, with the latter much more frequent in the marshes (6.2% vs 1.7%). By contrast, Hg T displayed a lower frequency in the marshes (7.6% vs 12.6%) due to a significant lower incidence of its T2 sub-clade (2.1% vs 6.9%, P < 0.05). On the other hand, Hgs N1 (8.2%) and W (4.8%), were both present in the marshes at a three-fold higher frequency than in Iraqis. Haplogroup X was detected as X2 with a frequency lower than 2% in both population samples.” ref
African haplogroups are of North/East and sub-Saharan African origin and represent minor components in both groups. The North/East African contribution is mainly represented by Hg M1 which accounts for 2.8% of Marsh Arabs and 1.2% of the Iraqi sample, the latter displaying also 0.6% of Hg U6. The sub-Saharan African component comprised Hgs L0, L1, L2 and L3 and accounted for 4.9% in the marshes and 9.1% of the control sample. Out of the twelve African sub-haplogroups identified in this survey, six in the marshes and seven in the control sample, only one (L2a1) was shared between the two Iraqi groups.” ref
“The Asian contribution was significantly higher (P < 0.01) in the Marsh Arabs than in the control sample (11.8% vs 5.2%). It includes mtDNAs belonging to the Southern Asian Hgs M (M*, M33, M37e) and R2 in Marsh Arabs, and R5a and U2d in the control sample. Haplogroup U7, frequent in Southwest Asia, was observed in both groups. The East Asian haplogroup B4 was detected at a very low frequency in both Iraqi groups.” ref
“Only a small proportion of the Marsh Arab gene pool derives from gene flow from neighboring regions. On the paternal side, our phylogeographic data highlight some southwest Asian specific contributions as testified to by Hgs Q, L, and R2, known as South Asian Y-chromosome lineages, primarily observed in India and Pakistan. Different from the Iraqi control sample, the Marsh Arab gene pool displays a very scarce input from the northern Middle East (Hgs J2-M172 and derivatives, G-M201 and E-M123), virtually lacks western Eurasian (Hgs R1-M17, R1-M412 and R1-L23) and sub-Saharan African (Hg E-M2) contributions. On the other hand, the absence in both Iraqi groups of the North African E-M81 branch, speaks against substantial patrilineal gene flow from this region.” ref
“On the maternal side, a significant (East/Southwest) Asian component (11.8%) is present among Marsh Arabs as testified to by Hgs B4, M, R2, and U7. The B4 mtDNAs carry control-region motifs observed in Iran, Kirghizstan, Western Siberia, Vietnam, and Korea, attesting to contact with Central and East Asia. This observation is likely due to recent gene flow, although it is worth noting that the ancient Silk Road passed through the Iraqi region from Basra to Baghdad. On the other hand, the majority of M, R2, and U7 mtDNAs display control-region motifs observed in South West Asia and, in particular, in India. Additional evidence of the multiple relationships with South West Asia derives from the presence of one M33 mtDNA, which was completely sequenced, (GenBank accession number: JN540042). This mtDNA belongs to the M33a2a clade and clusters with three sequences from Uttar Pradesh, Saudi Arabia, and Egypt, respectively. On the other hand, the presence in Iraq of Hgs M1 (in both Iraqi groups) and U6 (in the control sample) of North/East African origin is indicative of some limited gene flow from that area. The sub-Saharan contribution is instead represented by haplogroups L0, L1, L2, and L3. It reaches values (~8%) in line with those reported for other Middle Eastern Arab populations.” ref
“The last point is well illustrated by Y-chromosome data where the less represented J1-M267* lineage indicates Northern Mesopotamia contributions, whereas the most frequent J1-Page08 branch reveals a local recent expansion about 4,000 years ago. Although the Y-chromosome age estimates deserve caution, particularly when samples are small and standard errors large, it is interesting to note that these estimates overlap the City State period, which characterized Southern Mesopotamia and is testified to by numerous ancient Sumerian cities (Lagash, Ur, Uruk, Eridu, and Larsa).” ref
“In conclusion, our data show that the modern Marsh Arabs of Iraq harbor mtDNAs and Y chromosomes that are predominantly of Middle Eastern origin. Therefore, certain cultural features of the area, such as water buffalo breeding and rice farming, which were most likely introduced from the Indian sub-continent, only marginally affected the gene pool of the autochthonous people of the region. Moreover, a Middle Eastern ancestral origin of the modern population of the marshes of southern Iraq implies that, if the Marsh Arabs are descendants of the ancient Sumerians, also Sumerians were not of Indian or Southern Asian ancestry.” ref
“The Ma’dan, or Marsh Arabs, is a distinct group of people who originally inhabited the marshy area at the junction of the Tigris and Euphrates rivers in Iraq before the marshes were destroyed by irrigation projects developed by the Iraqi government. A seminomadic tribal people, the Ma’dan once lived in reed huts built on floating islands of reeds, and made their living by herding water buffalo, fishing, and hunting wild boars and waterfowl. Their houses were elaborately woven with Gothic-like arches made of bundles of reeds tied together at the top. This same kind of house had been built since the 4th millennium BCE.” ref
“The marshes were covered with rushes and reeds. Qasab, a kind of giant grass that looks like bamboo, covered most of the land and grew as tall as 7.6 m (25 ft). Natural islands, some floating and some anchored, dotted the waters, and the marshes were alive with wildlife, including turtles, frogs, various waterfowl, wild boars, and hordes of mosquitoes in the summer. Eagles were a common sight, soaring in the skies above the marshes. Summers were hot and humid; in the winter, the water was icy, and the winds were cold. A strong wind, called the “forty days’ wind,” blew throughout the month of June. Some historians believe the marshes were the Garden of Eden from the Bible.” ref
“The traditional Ma’dan believed in jinn, bad spirits who could take the form of humans or other animals. Unique to Ma’dan folklore are two marsh monsters: the anfish, a giant serpent with hairy skin, and the afa, a giant serpent with legs. Both were said to live somewhere in the heart of the marshes, and both were deadly. The Ma’dan also believed in a place called Hufaidh, an island of paradise located in the southwest part of the marshes, although no one knows exactly where. According to legends, the jinn could hide the island from human sight. On this island were palaces, palm trees, pomegranate orchards, and huge water buffalo. It was believed that anyone who saw Hufaidh was bewitched, and no one would be able to understand the person’s words afterward. Ma’dan boys are traditionally circumcised at puberty, but many boys refuse because of the frequent occurrence of infection afterward. After a death, some Ma’dan dye their turbans dark blue to signify mourning. Others put mud on their heads and clothes.” ref
“The traditional Ma’dan way of life was organized as a tribal society made up of various groups of families who shared a common lineage, with each family group headed by a sheikh (leader). Marriages were arranged by parents, although a couple had some choice in the matter. Paternal first cousins had the first claim to a young woman for their bride. Another who wished to marry her must have her paternal first cousin’s father’s agreement to give up his son’s right to her. In traditional Ma’dan homes, men and women did not eat together and all meals were conducted in silence. All talking was done before and after the meal, never during it. Men and women were generally segregated in public life as well, although young children would play together.” ref
“The displacement of the Ma’dan from their homeland in the marshes has caused the traditional tribal structure to deteriorate. Little is known about the conditions of Ma’dan women. Nearly 4.2 million Iraqis have fled the war-torn country, and an additional 2.2 million have left homelands within the country for cities and urban areas. The Ma’dan are among this group of displaced individuals, and until the situation in Iraq can be stabilized, the survival of both Ma’dan men and women is highly at risk.” ref
“One of the populations associated with haplogroup Q1b1 is the Marsh Arab population in the southern part of Mesopotamia. For thousands of years, this region has been a wetland due to its position between the Tigris and Euphrates rivers in modern-day Iraq. Mesopotamia has had human occupants since shortly after they first migrated from Africa 60,000-80,000 years ago. The current inhabitants of Mesopotamia, the Marsh Arabs, have strong ties to the ancient Sumer, one of the world’s first civilizations. Like their ancient forebears, Marsh Arab culture includes architecture based on arched reed buildings, subsistence strategies from grazing water buffalo, trapping birds, and spearing fish to cultivating rice, and the use of skinny boats called “Tarada” for transportation.” ref
“Haplogroup Q-L245 is linked to the Marsh ArabsOne of the populations associated with haplogroup Q1b1 is the Marsh Arab population in the southern part of Mesopotamia. For thousands of years, this region has been a wetland due to its position between the Tigris and Euphrates rivers in modern-day Iraq.” ref
Caucasus
“Caucasus hunter-gatherer (CHG), also called Satsurblia cluster, first identified in a 2015 study, based on the population genetics of several modern Western Eurasian (European, Caucasian, and Near Eastern) populations. It represents an ancestry maximized in some Upper Paleolithic and Mesolithic hunter-gatherer groups in the Caucasus. These groups are also very closely related to Mesolithic hunter-gatherers and Neolithic farmers in the Iranian Plateau, who are sometimes included within the CHG group. Ancestry that is closely related to CHG-Iranian Neolithic farmers is also known from further east, including from the Bactria–Margiana Archaeological Complex and the Harappan/Indus Valley Civilisation. Caucasus hunter-gatherers and Eastern hunter-gatherers are ancestral in roughly equal proportions to the Western Steppe Herders (WSH), who were widely spread across Europe and Asia beginning during the Chalcolithic.” ref
“The Mesolithic/Neolithic Iranian lineage basal to the Caucasus hunter-gatherers are inferred to derive significant amounts of their ancestry from Basal Eurasian (c. 38–48%), with the remainder ancestry being closer to Ancient North Eurasians or Eastern European Hunter-Gatherer (ANE/EHG; c. 52–62%). The CHG displayed an additional ANE-like component (c. 10%) than the Neolithic Iranians do, suggesting they may have stood in continuous contact with Eastern Hunter-Gatherers to their North. The CHG also carry around 20% additional Paleolithic Caucasus/Anatolian ancestry. Lazaridis et. al (2016) models the CHG as a mixture of Neolithic Iranians, Western Hunter-Gatherers and Eastern Hunter-Gatherers. In addition, CHG cluster with early Iranian farmers, who significantly do not share alleles with early Levantine farmers.” ref
“An alternative model without the need of significant amounts of ANE ancestry has been presented by Vallini et al. (2024), suggesting that the initial Iranian hunter-gatherer-like population which is basal to the CHG formed primarily from a deep Ancient West Eurasian lineage (‘WEC2’, c. 72%), and from varying degrees of Ancient East Eurasian (c. 10%) and Basal Eurasian (c. 18%) components. The Ancient West Eurasian component associated with Iranian hunter-gatherers (WEC2) is inferred to have diverged from the West Eurasian Core lineage (represented by Kostenki-14; WEC), with the WEC2 component staying in the region of the Iranian Plateau, while the proper WEC component expanded into Europe. Irving-Pease et. al (2024) models CHG as being derived from an Out of Africa population that split into basal Northern Europeans and West Asians. The latter was where CHG originated from.” ref
“At the beginning of the Neolithic, at c. 8000 BCE pr around 10,000 years ago, they were probably distributed across western Iran and the Caucasus, and people similar to the northern Caucasus and Iranian plateau hunter-gatherers arrived before 6000 BCE in Pakistan and north-west India. A roughly equal merger between the CHG and Eastern Hunter-Gatherers in the Pontic–Caspian steppe resulted in the formation of the Western Steppe Herders (WSHs). The WSHs formed the Yamnaya culture and subsequently expanded massively throughout Europe during the Late Neolithic and Early Bronze Age c. 3000—2000 BCE. Caucasus hunter gatherer/Iranian-like ancestry, was first reported as maximized in hunter-gatherers from the South Caucasus and early herders/farmers in northwestern Iran, particularly the Zagros, hence the label “CHG/Iranian.” ref
“Jones et al. (2015) analyzed genomes from males from western Georgia, in the Caucasus, from the Late Upper Palaeolithic (13,300 years old) and the Mesolithic (9,700 years old). These two males carried Y-DNA haplogroup: J* and J2a, later refined to J1-FT34521, and J2-Y12379*, and mitochondrial haplogroups of K3 and H13c, respectively. Their genomes showed that a continued mixture of the Caucasians with Middle Eastern populations took place up to 25,000 years ago, when the coldest period in the last Ice Age started. “CHG ancestry was also found in an Upper Palaeolithic specimen from Satsurblia cave (dated c. 11000 BCE), and in a Mesolithic one from Kotias Klde cave, in western Georgia (dated c. 7700 BCE).” ref
“The Satsurblia individual is closest to modern populations from the South Caucasus. Margaryan et al. (2017) analysing South Caucasian ancient mitochondrial DNA found a rapid increase of the population at the end of the Last Glacial Maximum, about 18,000 years ago. The same study also found continuity in descent in the maternal line for 8,000 years. According to Narasimhan et al. (2019) Iranian farmer related people arrived before 6000 BCE in Pakistan and north-west India, before the advent of farming in northern India. They suggest the possibility that this “Iranian farmer–related ancestry […] was [also] characteristic of northern Caucasus and Iranian plateau hunter-gatherers.” ref
“During the Neolithic and early Eneolithic, likely during the 4th millennium BC, Caucasus hunter-gatherers (CHGs) mixed with Eastern Hunter-gatherers (EHGs) on the Pontic–Caspian steppe, with the resulting population, almost half-EHG and half-CHG, forming the genetic cluster known as Western Steppe Herder (WSH). To David W. Anthony, Caucasus hunter-gatherer ancestry of Yamnaya is often with higher than 50%. As well as an overwhelming WSH ancestry, Yamnaya also have additional admixture from Anatolian and Levantine farmers, and the Western Hunter-gatherers (WHGs). Some scholars argue that the archaic PIE (‘Indo-Anatolian’) language may have originated among a CHG-rich population in Western Asia, based on the lack of EHG ancestry in the probable speakers of Anatolian languages. Others, such as Anthony, suggest that PIE was spoken by EHGs living in Eastern Europe.” ref
“According to Jones et al. (2015), Caucasus hunter-gatherer (CHG) “genomes significantly contributed to the Yamnaya steppe herders who migrated into Europe ~3,000 BCE, supporting a formative Caucasus influence on this important Early Bronze Age culture. CHG left their imprint on modern populations from the Caucasus and also Central and South Asia possibly correlating with the arrival of Indo-Aryan languages.” For example, about 50%-70% of Armenian ancestry is derived from CHG, persisting from Neolithic times to the present. Wang et al. (2018) analyzed genetic data of the North Caucasus of fossils dated between the 4th and 1st millennia BC and found a correlation with modern groups of the South Caucasus, concluding that “unlike today – the Caucasus acted as a bridge rather than an insurmountable barrier to human movement.” According to Allentoft et al. (2024), The arrival and admixture of CHG with Caspian steppe cultures is dated to about 7,300-years-old, which is seen in two ancient samples from Golubaya Krinitsa with 18-24% admixture.” ref
“CHG/Iranian Plateau Neolithic-like ancestry is prominent in pre-steppe admixture Chalcolithic and Bronze Age (4500–2000 BCE) populations in Central Asia, like the Bactria–Margiana Archaeological Complex (which also had Anatolian Neolithic Farmer-related ancestry) as well as in the northwestern Indian subcontinent such as in sites in or adjacent to the Indus Valley Civilisation (who have mixed CHG-related and Ancient Ancestral South Indian ancestry). It is unclear as to whether the dispersal of CHG/Iranian Plateau-related ancestry eastwards to the Indian subcontinent was the result of the migration of farmers or an earlier dispersal of hunter-gatherers who later adopted farming, but this dispersal likely occurred sometime before 6000 BCE due to the lack of Anatolian Farmer-related ancestry in ancient South Asians, but which is present in the Iranian Plateau after this time. This pre-steppe CHG-related ancestry makes up a significiant proportion of the ancestry of modern South Asians. WSHs, who were of significant CHG ancestry, also later migrated into Central Asia and the Indian subcontinent.” ref
“The 1100-kilometre long Caucasus mountain ranges extend between the Black Sea and the Caspian Sea and are bounded by the rivers Kuban and Terek in the north and the Kura and Araxes rivers in the south. The rich archaeological record suggests extensive human occupation since the Upper Palaeolithic. A Neolithic lifestyle based on food production began in the Caucasus after 6000 calBCE. As a region rich in natural resources such as ores, pastures, and timber, the Caucasus gained increasing importance to the economies of the growing urban centers in northern Mesopotamia. In the 4th millennium BCE, the archaeological record attests to the presence of the Maykop and Kura-Araxes, two major cultural complexes of the Bronze Age (BA) in the region. The Maykop culture is well known for its large and rich burial mounds, especially at the eponymous Maykop site, which reflects the rise of a new system of social organization, while the Kura-Araxes is found on both flanks of the Caucasus mountain range, demonstrating a connection between north and south.” ref
“Contact between the Near East, the Caucasus, the Steppe, and central Europe is documented, both archaeologically and genetically, as early as the 5th millennium BCE. This increased in the 4th millennium BCE along with the development of new technologies such as the wheel and wagon, copper alloys, new weaponry, and new breeds of domestic sheep. Such contact was critical in the cultural and genetic formation of the Yamnaya complex on the Eurasian Steppe—with about half of BA Steppe ancestry thought to derive from the Caucasus. In the 3rd millennium BCW, increased mobility associated with wheeled transport and the intensification of pastoralist practices led to dramatic expansions of populations closely related to the Yamnaya, accompanied by the domestication of horses allowing more efficient keeping of larger herds. These expansions ultimately contributed a substantial fraction to the ancestry of present-day Europe and South Asia. Thus, the Caucasus region played a crucial role in the prehistory and formation of Eurasian genetic diversity.” ref
“Recent ancient DNA studies have resolved several long-standing questions regarding cultural and population transformations in prehistory. One important feature is a cline of European hunter-gatherer (HG) ancestry that runs roughly from West to East. This ancestry differs from that of Early European farmers, who are more closely related to farmers of northwest Anatolia and also to pre-farming Levantine individuals. The Near East and Anatolia have long been seen as the regions from which European farming and animal husbandry emerged. In the Mesolithic and Early Neolithic, these regions harbored three divergent populations, with Anatolian and Levantine ancestry in the west and a group with a distinct ancestry in the east. The latter was first described in Upper Pleistocene individuals from Georgia (Caucasus hunter-gatherers; CHG) and then in Mesolithic and Neolithic individuals from Iran. The following millennia, spanning the Neolithic to BA, saw admixture between these ancestral groups, leading to a pattern of genetic homogenization of the source populations. North of the Caucasus, Eneolithic, and BA individuals from the Samara region (5200–4000 BCE) carry an equal mixture of EHG- and CHG/Iranian ancestry, so-called ‘steppe ancestry’ that eventually spread further west, where it contributed substantially to present-day Europeans, and east to the Altai region as well as to South Asia.” ref
“In our study, we aimed to investigate when and how the genetic patterns observed today were formed and test whether they have been present since prehistoric times by generating time-stamped human genome-wide data. We were also interested in characterizing the role of the Caucasus as a conduit for gene-flow in the past and in shaping the cultural and genetic makeup of the wider region. This has important implications for understanding the means by which Europe, the Eurasian steppe zone, and the earliest urban centers in the Near East were connected6. We aimed to genetically characterize individuals from cultural complexes such as the Maykop and Kura-Araxes and assessing the amount of gene flow in the Caucasus during times when the exploitation of resources of the steppe environment intensified, since this was potentially triggered by the cultural and technological innovations of the Late Chalcolithic and EBA around 4000–3000 BCE.” ref
“Finally, since the spread of steppe ancestry into central Europe and the eastern steppes during the early 3rd millennium BCE was a striking migratory event in human prehistory, we also retraced the formation of the steppe ancestry profile and tested for influences from neighboring farming groups to the west or early urbanization centers further south. Here we show that individuals from our Caucasian time transect form two distinct genetic clusters that were stable over 3000 years and correspond with eco-geographic zones of the steppe and mountain regions. This finding is different from the situation today, where the Caucasus mountains separate northern from southern Caucasus populations. However, during the early BA we also observe subtle gene flow from the Caucasus as well as the eastern European farming groups into the steppe region, which predates the massive expansion of the steppe pastoralists that followed in the 3rd millennium BCE.” ref
“The two distinct clusters are already visible in the oldest individuals of our temporal transect, dated to the Eneolithic period (~6300–6100 yBP/4300–4100 calBCE). Three individuals from the sites of Progress 2 and Vonyuchka 1 in the North Caucasus piedmont steppe (‘Eneolithic steppe’), which harbor EHG and CHG-related ancestry, are genetically very similar to Eneolithic individuals from Khvalynsk II and the Samara region. This extends the cline of dilution of EHG ancestry via CHG-related ancestry to sites immediately north of the Caucasus foothills. In contrast, the oldest individuals from the northern mountain flank itself, which are three first-degree-related individuals from the Unakozovskaya cave associated with the Darkveti-Meshoko Eneolithic culture (analysis label ‘Eneolithic Caucasus’), show mixed ancestry mostly derived from sources related to the Anatolian Neolithic (orange) and CHG/Iran Neolithic (green) in the ADMIXTURE plot. While similar ancestry profiles have been reported for Anatolian and Armenian Chalcolithic and BA individuals, this result suggests the presence of this mixed ancestry north of the Caucasus as early as ~6500 years ago.” ref
Ancient North Eurasian ancestry in Steppe Maykop individuals
“Four individuals from mounds in the grass steppe zone, archaeologically associated with the ‘Steppe Maykop’ cultural complex, lack the Anatolian farmer-related (AF) component when compared to contemporaneous Maykop individuals from the foothills. Instead they carry a third and fourth ancestry component that is linked deeply to Upper Paleolithic Siberians (maximized in the individual Afontova Gora 3 (AG3) and Native Americans, respectively, and in modern-day North Asians, such as North Siberian Nganasan. To illustrate this affinity with ‘ancient North Eurasians’ (ANE), we also ran PCA with 147 Eurasian and 29 Native American populations. The latter represents a cline from ANE-rich steppe populations such as EHG, Eneolithic individuals, AG3, and Mal’ta 1 (MA1) to modern-day Native Americans at the opposite end. To formally test the excess of alleles shared with ANE/Native Americans we performed f4-statistics of the form f4(Mbuti, X; Steppe Maykop, Eneolithic steppe), which resulted in significantly positive Z-scores (Z >3) for AG3, MA1, EHG, Clovis, and Kennewick for the ancient populations and many present-day Native American populations.” ref
“Based on these observations, we used qpWave and qpAdm methods to model the number of ancestral sources contributing to the Steppe Maykop individuals and their relative ancestry coefficients. Simple two-way models of Steppe Maykop as an admixture of Eneolithic steppe, AG3, or Kennewick do not fit. However, we could successfully model Steppe Maykop ancestry as being derived from populations related to all three sources (p-value 0.371 for rank 2): Eneolithic steppe (63.5 ± 2.9%), AG3 (29.6 ± 3.4%) and Kennewick (6.9 ± 1.0%). We note that the Kennewick related signal is most likely driven by the East Eurasian part of Native American ancestry as the f4-statistics (Steppe_Maykop, Fitted Steppe_Maykop; Outgroup1, Outgroup2) show that the Steppe Maykop individuals share more alleles not only with Karitiana but also with Han Chinese.” ref
“Individuals from the North Caucasian steppe associated with the Yamnaya cultural formation (5300–4400 BP, 3300–2400 calBCE) appear genetically almost identical to previously reported Yamnaya individuals from Kalmykia immediately to the north, the middle Volga region, Ukraine, and to other BA individuals from the Eurasian steppes who share the characteristic ‘steppe ancestry’ profile as a mixture of EHG and CHG-related ancestry. These individuals form a tight cluster in PCA space and can be shown formally to be a mixture by significantly negative admixture f3-statistics of the form f3(EHG, CHG; target). This cluster also involves individuals of the North Caucasus culture (4800–4500 BP, 2800–2500 calBCE) in the piedmont steppe, who share the steppe ancestry profile, as do individuals from the Catacomb culture in the Kuban, Caspian, and piedmont steppes (4600–4200 BP, 2600–2200 calBCE), which succeeded the Yamnaya horizon.” ref
“The individuals of the MBA post-Catacomb horizon (4200–3700 BP, 2200–1700 calBCE) such as Late North Caucasus and Lola cultures represent both ancestry profiles common in the North Caucasus: individuals from the mountain site Kabardinka show a typical steppe ancestry profile, whereas individuals from the site Kudachurt 90 km to the west or our most recent individual from the western LBA Dolmen culture (3400–3200 BP, 1400–1200 calBCE) retain the ‘southern’ Caucasus profile. In contrast, one Lola culture individual resembles the ancestry profile of the Steppe Maykop individuals.” ref
“Evidence for interaction between the Caucasus and the Steppe clusters is visible in our genetic data from individuals associated with the later Steppe Maykop phase around 5300–5100 years ago. These ‘outlier’ individuals were buried in the same mounds as those with steppe and, in particular, Steppe Maykop ancestry profiles but share a higher proportion of AF ancestry visible in the ADMIXTURE plot and are also shifted towards the Caucasus cluster in PC space. This observation is confirmed by formal D-statistics. By modeling Steppe Maykop outliers successfully as a two-way mixture of Steppe Maykop and representatives of the Caucasus cluster, we can show that these individuals received additional ‘Anatolian and Iranian Neolithic ancestry,’ most likely from contemporaneous sources in the south. We used ALDER to estimate an average admixture time for the observed farmer-related ancestry in Steppe Maykop outliers of 20 generations or 560 years ago.” ref
“Eneolithic Samara individuals form a cline in PC space running from EHG to CHG, which is continued by the newly reported Eneolithic steppe individuals. However, the trajectory of this cline changes in the subsequent centuries. Here we observe a cline from Eneolithic_steppe towards the Caucasus cluster. We can qualitatively explain this ‘tilting cline’ by developments south of the Caucasus, where Iranian and AF ancestries continue to mix, resulting in a blend that is also observed in the Caucasus cluster, from where it could have spread onto the steppe. The first appearance of ‘combined farmer-related ancestry’ in the steppe zone is evident in Steppe Maykop outliers. However, PCA results suggest that Yamnaya and later groups of the West Eurasian steppe carry also some farmer-related ancestry as they are slightly shifted towards ‘European Neolithic groups’ in PC2 compared to the preceding Eneolithic steppe individuals. The ‘tilting cline’ is also confirmed by admixture f3-statistics, which provide statistically significant negative values for AG3 and any AF group as the two sources. Using f– and D-statistics we also observe an increase in farmer-related ancestry (both Anatolian and Iranian) in our Steppe cluster, distinguishing the Eneolithic steppe from later groups. In addition, we find the Caucasus cluster or Levant/AF groups to share more alleles with Steppe groups than with EHG or Samara_Eneolithic. MLBA groups such as Poltavka, Andronovo, Srubnaya, and Sintashta show a further increase of AF ancestry consistent with previous studies, reflecting different processes not directly related to events in the Caucasus.” ref

R1a
“R1a could have migrated directly to eastern Europe (European Russia, Ukraine, Belarus), or first southward through Central Asia and Iran. In that latter scenario, R1a would have crossed the Caucasus during the Neolithic, alongside R1b, to colonize the Pontic-Caspian Steppe. In the absence of ancient Y-DNA from those regions, the best evidence supporting a Late Paleolithic migration to Iran is the presence of very old subclades of R1a (like M420) in the region, notably in the Zagros mountains. However, these samples only make up a fraction of all R1a in the region and could just as well represent the descendants of Eastern European hunter-gatherers who branched off from other R1a tribes and crossed from the North Caucasus any time between 20,000 and 8,000 years ago. The logic behind this is that most known historical migrations in Eurasia took place from north to south, as people sought warmer climes. The only exception happened during the Holocene warming up of the climate, which corresponds to the Neolithic colonization of Europe from the Near East. A third possibility is that R1a tribes split in two around Kazakhstan during the Late Paleolithic, with one group moving to eastern Europe, while the other moved south to Iran.” ref
“Some people have theorized that R1a was one of the lineages of the Neolithic farmers, and would have entered Europe through Anatolia, then spread across the Balkans toward Central Europe, then only to Eastern Europe. There are many issues with this scenario. The first is that 99% of modern R1a descends from the branch R1a-M417, which clearly expanded from the Bronze Age onwards, not from the early Neolithic. Its phylogeny also points at an Eastern European origin. Secondly, most of the R1a in the Middle East are deep subclades of the R1a-Z93 branch, which originated in Russia (see below). It could not have been ancestral to Balkanic or Central European R1a. Thirdly, there is a very strong correlation between the Northeast European autosomal admixture and R1a populations, and this component is missing from the genome of all European Neolithic farmers tested to date – even from Ötzi, who was a Chalcolithic farmer. This admixture is also missing from modern Sardinians, who are mostly descended from Neolithic farmers. This is incontrovertible evidence that R1a did not come to Europe with Neolithic farmers, but only propagated from Eastern Europe to the rest of Europe from the Bronze Age onwards.” ref
“R1a is thought to have been the dominant haplogroup among the northern and eastern Proto-Indo-European tribes, which evolved into the Indo-Iranian, Thracian, Baltic, and Slavic people. The Proto-Indo-Europeans originated in the Yamna culture (3300-2500 BCE). Their dramatic expansion was possible thanks to an early adoption of bronze weapons and the domestication of the horse in the Eurasian steppes (circa 4000-3500 BCE). Individuals from the southern part of the Steppe are believed to have carried predominantly lineages belonging to haplogroup R1b (L23 and subclades), while the people of northern forest-steppe to the north would have belonged essentially to haplogroup R1a. The first expansion of the forest-steppe people occurred with the Corded Ware Culture (see Germanic branch below).” ref
“The forest-steppe origin of this culture is obvious from the usage of corded pottery and the abundant use of polished battle axes, the two most prominent features of the Corded Ware culture. This is also probably the time when the satemisation process of the Indo-European languages began, considering that the Balto-Slavic and Indo-Iranian language groups belong to the same Satem isogloss, and both appear to have evolved from the Catacomb and Srubna cultures. Ancient DNA testing has confirmed the presence of haplogroup R1a-M417 in samples from the Corded Ware culture in Germany (2600 BCE), from Tocharian mummies (2000 BCE) in Northwest China, from Kurgan burials (circa 1600 BCE) from the Andronovo culture in southern Russia and southern Siberia, as well as from a variety of Iron-age sites from Russia, Siberia, Mongolia and Central Asia.” ref
“The origins of the Slavs go back to circa 3500 BCE with the northern Yamna culture and its expansion across Central and Northeast Europe with the Corded Ware culture. The M458 and Z280 lineages spread around Poland, Belarus, Ukraine, and western Russia and would form the core of the Proto-Balto-Slavic culture. The high prevalence of R1a in Baltic and Slavic countries nowadays is not only due to the Corded Ware expansion, but also to a long succession of later migrations from Russia, the last of which took place from the 5th to the 10th century CE. The Slavic branch differentiated itself when the Corded Ware culture absorbed the Cucuteni-Tripolye culture (5200-2600 BCE) of western Ukraine and north-eastern Romania, which appears to have been composed primarily of G2a-U1 et I2a1b-M423 lineages descended directly from Paleolithic Europeans, with some other Near-Eastern farmer lineages (notably E-V13, J2a and T1a).” ref
“It is surely during this period that I2a2, E-V13, and T spread (along with R1a) around Poland, Belarus, and western Russia, explaining why eastern and northern Slavs (and Lithuanians) have between 10 and 20% of I2a1b lineages and about 10% of Middle Eastern lineages (18% for Ukrainians). The Corded Ware period was followed in the steppes by the Srubna culture (1800-1200 BCE), and around Poland by the Trzciniec culture (1700-1200 BCE). Mathieson et al. (2015) and Krzewinska et al. (2018) each retrieved the Y-DNA from six Srubna individuals, and all 12 of them belonged to haplogroup R1a. Those tested for deep clades were positive for Z93, the Indo-Iranian branch.” ref
From the Epipalaeolithic into the earliest Neolithic (PPNA) in the South Levant
“Abstract: This paper examines the nature of initial neolithisation indications during the terminal Pleistocene and earliest Holocene in the Southern Levant. This interval corresponds to a period of significant and geographically variable environmental changes in the region. Various lines of evidence are provided to demonstrate the long durée (~15 000 years) character of interactions during the Early, Middle, and Late Epipalaeolithic that was instrumental to the emergence of the fully-fledged agricultural lifeways in the later phases of the Early Neolithic (PPNB).” ref
“Since the inception of research in Southwest Asia, the unique ‘bridging’ role of the Natufian culture, between the ‘Palaeolithic’ and the ‘Neolithic’ (with all that such a role implies) has been widely acknowledged. The ‘intermediate’ nature of this entity – representing the shift from the preceding groups of mobile hunter-gatherers towards the following settled farming societies, especially in the Mediterranean zone – is reflected in many aspects of its material culture and lifeways. There is, for the first time, evidence of long-term, semi-sedentary basecamp sites with substantial, durable structures accompanied by a largely microlithic knapped industry (Palaeolithic in nature), together with a heavy-duty component and sickle blades (heralding the chipped stone traditions of the Neolithic).” ref
“Other distinctive features include cemeteries exhibiting complex burial rites, extensive groundstone and bone tool assemblages, and the appearance of artistic and symbolic manifestations as constant components of the material culture repertoire. Yet, after many years of systematic research, the terminology and processes involved in the transformation from the latest Natufian to the first Neolithic entities, namely the Pre-Pottery Neolithic A (PPNA; incorporating at least two cultural entities, the ‘Khiamian’ and the ‘Sultanian’) has been contentious and hotly debated. Additionally, there has been increasing awareness that, at least in the Southern Levant, the criteria that justified the use of the term ‘Neolithic Revolution’ actually became apparent only with the shift from PPNA to PPNB.” ref
“Initially, as is often the case in archaeology, it was assumed that the Neolithic ‘package’ was ‘exotic’ (i.e. allochthonous), having arrived from somewhere outside the region, as it was assumed that major changes as a rule occurred through external ‘stimuli’. Later, it was believed that during the latest Natufian phase there was a reversion to a more mobile life-ways, brought about mostly due to the ‘forcing’ conditions of the supposedly harsh climatic Younger Dryas event. Still, it was assumed that the local Neolithic rose from the ‘ashes’ of the declining Natufian, without going into the specifics of how this came about, or the relation of the local southern Neolithic to the emergence of early Neolithic phenomena in the northern Levant. That the local Neolithic represents Natufian survival, ‘by-the-skin-of-their-teeth’, is strengthened also by the presence of only a few sites with a unbroken Natufian – Neolithic sequence and the fact that most PPNA occurrences are found in different locations from the preceding major Natufian base camps.” ref
“Current research has clearly demonstrated that there is solid evidence for local, in situ continuity between the two cultural manifestations. This is reflected by recent investigations of several ‘Epi-Natufian’ and ‘Khiamian’ settlements in the Mediterranean zone. Sites are mostly located at low elevations at the edge of the lower Jordan valley, i.e. Salibiya IX, Gilgal II, Huruk Musa and Nahal Ein Gev II and in the low-lying areas west of the central hill range, i.e. Nahal Oren, Tel Bareqet, Tzur Nathan, Kaizer, Qula and Hatoula, all dating to the interval of c. 12,500–11,600 cal years ago. The archaeology of those sites provides solid evidence of local, endemic developments ‘bridging’ the end of the Natufian complex foragers’ existence and the emergence of larger, clearly sedentary PPNA (Sultanian) settlements. In attempting to understand the dynamics that operated to enable the appearance of what is considered to represent the initial Neolithic, it is necessary to refer to the Early and Middle Epipalaeolithic. Indeed, quite a number of ‘Natufian-cum-Neolithic’ characteristics appear much earlier, already by the Early Epipalaeolithic, with the onset of the Last Glacial Maximum.” ref
“This said, we shall present rather briefly the processes believed to be instrumental in shaping the Natufian entity, and which apparently continued into the Neolithic. Clearly, some of what will be presented is rather speculative, but we do rely on evidence to that effect in the archaeological record. We should clarify that we adhere to and point out the separation between long-term and short-term trends taking place throughout the entire Epipalaeolithic sequence.” ref
“Something old, something new, something borrowed, something blue…” Ornaments in the Levantine Early Neolithic
“Abstract: With the onset of the Near Eastern Neolithic during the 12th millennium cal years ago or 12,000 to 11,000 years ago, and thereafter, one can observe growing sedentary tendencies, as well a significant increase in populations and community sizes, all reflected in the Neolithic demographic transition. At that time (and even somewhat earlier in certain areas) a notable tendency for within and between community differentiation was observed, archaeologically visible through the variances in the material remains. A specific domain where this phenomenon can be observed are the easily portable items of adornment. The aspiration for symbolling and signaling at both the community level and the individual served to increase webs of interactions and exchange between communities, sometimes over huge distances. The differences and the similarities actually reflected the degree and intensity of connectivity between the communities far and wide.” ref
NATUFIANS (12,500-9500 BCE): SETTLEMENTS, PROTO-AGRICULTURE
“The Natufian culture refers to mostly hunter-gatherers who lived in modern-day Israel, Jordan, Lebanon, and Syria approximately 15,000 to 11,500 years ago. Merging nomadic and settled lifestyles, they were among the first people to build permanent houses and cultivate edible plants. The advancements they achieved are believed to have been crucial to the development of agriculture during the time periods that followed them. According to Encyclopaedia Britannica: Mainly hunters, the Natufians supplemented their diet by gathering wild grain; they likely did not cultivate it. They had sickles of flint blades set in straight bone handles for harvesting grain and stone mortars and pestles for grinding it. Some groups lived in caves, others occupied incipient villages. They buried their dead with their personal ornaments in cemeteries. Carved bone and stone artwork have been found.” ref
“Matti Friedman wrote in Smithsonian magazine: Natufians were linked by characteristic tools, particularly a small, half-moon-shaped flint blade called a lunate,. They also showed signs of the “intentional cultivation” of plants, according to Ofer Bar-Yosef, professor of prehistoric archaeology at Harvard University, using a phrase that seems carefully chosen to avoid the loaded term “agriculture.” Other characteristic markers included jewelry made of dentalium shells, brought from the Mediterranean or the Red Sea; necklaces of beads made of exquisitely carved bone; and common genetic characteristics like a missing third molar. A study, published in Nature Scientific Reports by a team of scientists and archeologists from the Weizmann Institute of Science in Rechovot and the University of Copenhagen, rejects the long-held “core region” theory that argues that the Natufian culture spread from the Mount Carmel and Galilee region and suggest instead the Natufians had far more diverse and complex origins.” ref
“Daniel K. Eisenbud wrote in the Jerusalem Post: “According to the researchers, the study is based on evidence from a Natufian site located in Jordan, some 150 km. northeast of Amman. The site, called Shubayqa 1, was excavated by a University of Copenhagen team led by Dr. Tobias Richter from 2012-2015. The excavations uncovered a well-preserved Natufian site, which included, among other findings, a large assemblage of charred plant remains. The botanical remains, which are rare in many Natufian sites in the region, enabled the Weizmann-Copenhagen team to obtain the largest number of dates for any Natufian site yet in either Israel or Jordan. Utilizing an accelerator mass spectrometer (AMS), that can reveal the amount of carbon-14 in a sample as small as a single atom, Prof. Elisabetta Boaretto of the Weizmann Institute, was able to accurately date the charred remains. “To ensure the highest accuracy, the team selected only samples from short-lived plant species or their parts – for example, seeds or twigs – to obtain the dates. “Over 20 samples from different layers of the site were dated, making it one of the best and most accurately dated Natufian sites anywhere,” the Institute continued. “The dates showed, among other things, that the site was first settled not long after the earliest dates obtained for northern Israel.” ref
“Based on the findings, the researchers concluded that either Natufians spread very rapidly into the region, or, more probably, that the settlement patterns emerged more or less simultaneously in different parts of the region. “The early date of Shubayqa 1 shows that Natufian hunter- gatherers were more versatile than previously thought,” said Richter. “Past research had linked the emergence of Natufian culture to the rich habitat of the Mediterranean woodland zone. But the early dates from Shubayqa 1 show that these late Pleistocene hunter-gatherers were also able to live quite comfortably in more open parkland steppe zones further east.” Richter noted the researchers determined that a portion of the Natufians’ subsistence appears to have relied heavily on the exploitation of club rush tubers as well as other wild plants and the hunting of birds, gazelle, and other animals. According to Boaretto, the “core area” theory may have come about, in part, because the Mount Carmel sites have been the best preserved and studied – until now. “In addition to calling into question the idea that the Natufians originated in one settlement and spread outwards, the study suggests that the hunter- gatherers who lived 15,000 to 12,000 years ago were ingenious and resourceful,” said Boaretto. The authors concluded that their findings support the view that there were many pathways to agriculture and “the Neolithic way of life” was a highly variable and complex process that cannot be explained on the basis of single-cause models.” ref
“Laura Anne Tedesco of the Metropolitan Museum of Art wrote: “The Natufians were the first people of the eastern Mediterranean area to establish permanent villages. Prior to the Natufians, bands of people had moved seasonally, to follow animals for hunting and to gather available plants. The Natufians, while still hunters and foragers, settled in villages year-round, relying on the natural resources of their immediate area. These resources included gazelle, wild cereals, and marine life. The latter, abundant in the region, was used for food as well as for making tools, art, and body ornamentation. Shells collected from the Mediterranean and the Red Sea were commonly used for jewelry and headdresses, typical status markers. “After the last Ice Age, as the climate became warmer and rainfall more abundant, the nomadic population of the eastern Mediterranean began to establish the first permanent settlements. The site of Eynan/Ain Mallaha, situated between the hills of Galilee and Lake Hula in the Levant, was inhabited from 10,000 to 8,200 BCE or around 12,000-10,200 years ago, during the Natufian period. Eynan (in Hebrew)/Ain Mallaha (in Arabic) is one of the hundreds of Natufian settlements known from the eastern Mediterranean, where remains of a rich and dynamic artistic tradition have been discovered.” ref
“Excavations in the Levant, including at Eynan/Ain Mallaha, were undertaken with great enthusiasm by European and American archaeologists in the years following World War II. During this period of scientific exploration, hundreds of sites were uncovered, not just Natufian but from preceding and succeeding periods. These archaeological activities contributed enormously to our current understanding of the prehistoric record of this region. Jericho, well known for its defensive walls described in biblical accounts, is another important Natufian site that was discovered at about the same time as Eynan/Ain Mallaha.” “When we talk about people in the Natufian period becoming more sedentary and less mobile, it’s not just about economics and settlement patterns, but about culture,” Tobias Richter, a professor of prehistory at the University of Copenhagen, told Smithsonian. “At what point does a place become home? At what point do we develop emotions and attachments that are tied to a specific location?” ref
“Natufian settlements were generally found in open woodland areas with oak and Pistacia trees and underbrush with large amounts of grain-carrying grasses. They tended to stay away from the high mountains of Lebanon, the steppe areas of the Negev desert in Israel and Sinai, and the Syro-Arabian desert in the east, presumably because of limited food resources and competition from other groups of foragers who exploited this region. Natufian dwellings were semi-subterranean, often with a dry-stone foundation. The frame was probably made of brushwood. No traces of mudbrick have been found, which became common later in the Pre-Pottery Neolithic A (PPNA) period. Their round houses have a diameter between three and six meters, and contain a central round or subrectangular fireplace. In Ain Mallaha traces of postholes have been identified. Villages covering over 1,000 square meters have been found. Smaller settlements have been interpreted by some researchers as camps. Traces of rebuilding in almost all excavated settlements seem to point to a frequent relocation, hinting of a semi-nomadic rather than settled existence. Settlements have been estimated to house 100–150 people, but there are three categories: small, medium, and large, ranging from 15 square meters to 1,000 square meters. There are no definite indications of storage facilities.” ref
“The Natufian people lived by hunting and gathering. The preservation of plant remains at Natufian sites is poor because of the soil conditions, but wild cereals, legumes, almonds, acorns, and pistachios may have been collected. Animal bones show that gazelle (Gazella gazella and Gazella subgutturosa) were the main prey. Deer, aurochs and wild boar were hunted in the steppe zone, as well as onagers and caprids (ibex). Water fowl and freshwater fish formed part of the diet in the Jordan River valley. Animal bones from Salibiya have been interpreted as evidence for communal hunts with nets. A pita-like bread dated to 12.500 BCE or around 14,500 years ago has been attributed to Natufians. This bread is made of wild cereal seeds and papyrus cousin tubers, ground into flour.” ref
“According to one theory, a sudden change in climate — the Younger Dryas event (c. 10,800 to 9500 BCE) — inspired the development of agriculture. The Younger Dryas was a 1,000-year-long interruption in the higher temperatures prevailing since the Last Glacial Maximum, which produced a sudden drought in the Levant. This could have threatened wild cereals, which were out-competed by dryland scrub. It is presumed that local population had become largely sedentary. To preserve their sedentary way of life they cleared the scrub and planting seeds obtained from elsewhere, originating agriculture. This theory is controversial and hotly debated in the scientific community. Ancient figs found in an archaeological site in the Jordan Valley, presumably where Natufian lived , may represent one of the earliest forms of agriculture, scientists report. The carbonised fruits date between 11,200 and 11,400 years old. The U.S. and Israeli researchers reported their findings in the journal Science and said the figs are a variety that could have only been grown with human intervention, arging the discovery marks the point when humans turned from hunting and gathering to food cultivation.” ref
“Rebecca Morelle of the BBC reported: “Nine small figs, measuring just 18mm (0.7in) across, along with 313 smaller fig fragments were discovered in a house in an early Neolithic village, called Gilgal I, in the Jordan Valley. The researchers from Harvard University in and Bar-Ilan University in Israel believe the figs are an early domestic crop rather than a wild breed. The ancient fig is smaller than these varieties of modern fig After examining the figs, they determined that it was a self-pollinating, or parthenocarpic, variety, like the kind we eat today. In nature, parthenocarpic fig trees appear now and again by a chance genetic mutation; but because they do not produce seeds, they cannot reproduce alone — they require a shoot to be removed and replanted.” ref
Natufians and the Neolithic Revolution?
“V. Gordon Childe (1892-1957, an Australian transplant to Britain, invented the concept of the Neolithic Revolution in the 1920s. Charles C. Mann wrote in National Geographic, ” In today’s terms, Childe’s views could be summed up like this: Homo sapiens burst onto the scene about 200,000 years ago. For most of the millennia that followed, the species changed remarkably little, with humans living as small bands of wandering foragers. Then came the Neolithic Revolution — “a radical change,” Childe said, “fraught with revolutionary consequences for the whole species.” In a lightning bolt of inspiration, one part of humankind turned its back on foraging and embraced agriculture. The adoption of farming, Childe argued, brought with it further transformations. To tend their fields, people had to stop wandering and move into permanent villages, where they developed new tools and created pottery. The Neolithic Revolution, in his view, was an explosively important event — “the greatest in human history after the mastery of fire.” ref
“Matti Friedman wrote in Smithsonian magazine: The discoveries at Ein Gev challenge the conventional wisdom about the agricultural revolution—and raise the question of whether the term “revolution” is even the right one… Childe was deeply affected by the Industrial Revolution, which had altered the Western world by the time of his birth, and the 1917 Bolshevik Revolution in Russia, which happened when he was 25. In the first half of the 20th century, which saw the breakup of old empires and ascendant movements for individual rights, belief in scientific progress and human agency was deeply felt. The promise of Childe’s “agricultural revolution” was a life freed from the unpredictable labor of hunting and gathering, and a step up the ladder toward an ordered way of life.” ref
“Today, many scholars still identify the move to agriculture as a singular moment of human invention that paved the way for modern life. This is supported by genetic studies and archaeological finds that trace the appearance of domesticated grains to modern-day Turkey and Syria approximately 10,000 years ago. Avi Gopher, an emeritus professor of archaeology at Tel Aviv University, for example, pinpoints the transformation to 10,500 years ago. He calls this a “big bang”—a leap in plant domestication in southeastern Turkey executed not gradually but rapidly and purposefully, based on what we would think of as scientific understanding. Smart people, in other words, figured out something novel and important, and human history was never the same after that. This breakthrough, he told me, can only be called a revolution. “The idea that you could take control of a species in nature and put it to work for you—this was a complete change of worldview, one that led, in many ways, to our own civilization. And the evidence is that it happened quickly.” ref
“The key moment of the “agricultural revolution” is sometimes described as a lightning bolt of human innovation akin to electricity or powered flight. It’s a good story. But at Nahal Ein Gev II, every dig season complicates it a little more. The village here flourished 2,000 years before the revolution got going, and the growing impression is of a place curiously ahead of its time—“a true turning point in human culture,” in the words of Steven Mithen, the eminent British prehistorian and author of After the Ice: A Global Human History. Matti Friedman wrote in Smithsonian magazine: The finds at Nahal Ein Gev II testify to humans settling down and practicing some form of cultivation millennia before that “big bang.” “You’ll never hear me say the word ‘revolution,’” Grosman told me. “I hate the word. If we look at Ein Gev, we’ll see that exciting things are happening there, but it’s not a ‘revolution.’ They’ve got one foot in one era and the other foot in another, and that’s why they’re so interesting.” ref
“It’s not that Grosman disagrees that people figured out how to harness grain mutations in southeastern Turkey 2,000 years later. But she sees that as a late stage of the process, not the beginning, and as more of an elaboration than the breakthrough itself. The leap, in her eyes, was from nomadism to living in one place, harvesting plants, and building a society bigger than an extended family. This shift is visible at Nahal Ein Gev II, she believes, and this change made possible the invention of agriculture—not the other way around. The truth is that revolutions have always been more complicated than we tend to think. Was the French Revolution caused by the oppression of common people by royal autocrats, by the spread of liberal ideals, or by expanding literacy? Or was it due to the actions of specific people like Louis XVI or Robespierre? The answer is some combination that is impossible to predict at the time and hard to grasp afterward. Richter, the University of Copenhagen prehistorian, views the move to agriculture in the same way. “I think we have to see history and historic process as an almost chaotic overlapping of circumstances that co-occur at the same time,” he said.” ref
“The Khiamian Period (c. 10,200 – c. 8,800 BCE or around 12,200-10,800 years ago, also referred to as El Khiam or El-Khiam) is a period of the Near-Eastern Neolithic, marking the transition between the Natufian and the Pre-Pottery Neolithic. Some sources date it from about 10,000 to 9,500 BCE, The Khiamian is named after the site of El Khiam, situated on banks of the Dead Sea, where researchers have recovered the oldest chert arrows heads, with lateral notchs, the so-called “El Khiam points”, which has served to identify sites in Israel, (Azraq), Sinai (Abu Madi), and to the north as far as the Middle Euphrates (Mureybet). The Khiamian is regarded as a time without any major technical innovations. However, for the first time, houses were built on the ground level, not half buried as was previously done. Otherwise, members of this culture were still hunter-gatherers. Agriculture was still rather primitive. Relatively new discoveries in the Middle East and Anatolia show that some experiments with agriculture had taken place by 10,900 BCE, and wild grain processing had occurred by 19,000 BCE at Ohalo II. According to Jacques Cauvin, the Khiamian was the beginning of the worship of the Woman and the Bull, found in later following periods in the Near-East, based on the appearance of small female statuettes, as well as by the burying of aurochs skulls.” ref
“Not far from the Sea of Galilee is the Natufian site of Nahal Ein Gev II, which is sometimes described as a village. Matti Friedman wrote in Smithsonian magazine: I came to the village with Leore Grosman, an archaeologist from the Hebrew University of Jerusalem….Her team of 30 arrived in work pants and sun hats from their quarters at a nearby kibbutz. At first light, they fanned out across the hill by the stream. Soon a few were squatting in the remains of a round house. Several diggers under the direction of Natalie Munro, an archaeologist from the University of Connecticut, were busy in the adjacent cemetery, brushing off an adult cranium and treading carefully around the skeleton of a 3-year-old. One team member set up a geolocation tripod that precisely locates every artifact on a grid. A PhD student looked for gazelle bones. The pace picked up as the sun rose, the same atmosphere of industry you might have sensed if you had come when the villagers were here 12 millennia ago.” ref
“The village yields new surprises each season. When I was there, a pair of doctoral students from Canada and Sweden were brushing earth from two reddish-brown objects inside one of the Natufian homes. It wasn’t clear what the objects were, but they were made of clay, which didn’t seem notable at first—anyone who has spent time at an archaeological dig has seen plenty of pottery. But then I remembered that pottery wasn’t supposed to have been invented in this part of the world for another 4,000 years. “The finds at the site make clear that its people were innovators,” Mithen, the British prehistorian, told me. “Processes of change that we thought occurred during the later Neolithic were already underway at this settlement. Nahal Ein Gev illustrates how architecture, art and economy are interlinked in ways that we have yet to fully understand in the transition from hunting and gathering to farming lifestyles.” ref



")
“Haplogroup R2a (R-M124) is characterized by SNPs M124, F820/Page4, L381, P249, and is mainly found in South Asia, with lower frequencies in Central Asia. R-M124 is also found in multiple Jewish populations: Iraqi Jews, Persian Jews, Mountain Jews, and Ashkenazi Jews. Most research has tested only for the presence of R-M479 (R2) and R-M124 (R2a) – or SNPs downstream from M124 like P249, P267, L266, PAGES00004, and L381 SNPs). Because the other primary branch, R2b (R-FGC21706) was discovered later than R2a, it has often not been tested for. Hence most results are best described as R2(xR2a). n addition, relatively little research has been done within South Asia, which is known to have the greatest concentration of R2. (Hence the figures cited in the table right may not be indicative of true frequencies, i.e. Pakistan is the only South Asian country that has been included.) In 2013, R2(xR2a) was found in 5 out of 19 males from the Burusho minority of North Pakistan. Haplogroup R2, or R-M479, has been concentrated geographically in South Asia and Central Asia since prehistory. It appears to reach its highest levels among the Burusho people in North Pakistan.” ref
“Haplogroup R2a, or haplogroup R-M124, is a Y-chromosome haplogroup characterized by genetic markers M124, P249, P267, L266, and is mainly found in South Asia as well as in Central Asia, Caucasus, West Asia and North Africa / MENA. Haplogroup R-M124 was known as Haplogroup P1 and formerly thought to be a sister clade of Haplogroup R rather than derived from it. Haplogroup R2 most often observed in Asia, especially on the Indian sub-continent and Central Asia. It is also reported at notable frequencies in Caucasus.” ref
“Ancient samples of haplogroup R2a were observed in the remains of humans from Neolithic, Chalcolithic, and Bronze Age Iran and Turan; and Iron Age South Asia. R2a was also recovered from excavated remains in the South Asian sites of Saidu Sharif and Butkara from a later period. R-M124 is most often observed in Asia, especially on the Indian sub-continent and in Central Asia. It is also reported at notable frequencies in Caucasus. Haplogroup R-M124, along with haplogroups H, L, R1a1, and J2, forms the majority of the South Asian male population. The frequency is around 10-15% in India and Sri Lanka and 7-8% in Pakistan. Its spread within South Asia is very extensive, ranging from Baluchistan in the west to Bengal in the east; Hunza in the north to Sri Lanka in the south.” ref
“According to Sengupta et al. (2006), uncertainty neutralizes previous conclusions that the intrusion of HGs R1a1 and R2 [Now R-M124] from the northwest in Dravidian-speaking southern tribes is attributable to a single recent event. Rather, these HGs contain considerable demographic complexity, as implied by their high haplotype diversity. Specifically, they could have actually arrived in southern India from a southwestern Asian source region multiple times, with some episodes considerably earlier than others.” ref
| Haplogroup R‑M124 |
|
||||||||||||
Paragroup R-M124*
Haplogroup R-L295
“Haplogroup R-L295 is a Y-chromosome haplogroup characterized by genetic marker L295. It is found in South Asia, Anatolia, Arabian Peninsula, Europe, & Central Asia so far.” ref
Haplogroup R-L263
“Haplogroup R-L263 is a Y-chromosome haplogroup characterized by genetic marker L263. It is found in Greek Asia Minor & Armenia so far.” ref
Haplogroup R-L1069
“Haplogroup R-L1069 is a Y-chromosome haplogroup characterized by genetic marker L1069. It is found in Kuwait so far.” ref
“The haplogroup R-M124 frequency of 6.1% (6/114) was found among overall Kurds while in one study which was done with 25 samples of Kurmanji Kurds from Georgia, R-M124 has been observed at 44% (11/25). In Caucasus high frequency was observed in Armenians from Sason at 17% (18/104) while it was observed at %1 in Armenians from Van. R2 has been found in Chechens at 16%. R-M124 has been found in approximately 8% (2/24) of a sample of Ossetians from Alagir. In the Caucasus, around 16% of Mountain Jews, 8% of Balkarians, 6% of Kalmyks, 3% of Azerbaijanis, 2.6% of Kumyks, 2.4% of Avars, 2% of Armenians, and 1% to 6% of Georgians belong to the R-M124 haplogroup. Approximately 1% of Turks and 1% to 3% of Iranians also belong to this haplogroup.” ref
“In Iran R-M124 follows a similar distribution as R1a1 with higher percentages in the southeastern Iran. It has been found at Frequencies of 9.1% at Isfahan, 6.9% at Hormozgan, and 4.2% in Mazandaran. In the R2-M124-WTY and R-Arabia Y-DNA Projects, Haplogroup R-M124 has appeared in the following Arab countries: Kuwait (3 clusters), United Arab Emirates (1 cluster), Syrian Arab Republic (1 cluster), and Tunisia (1 cluster). Thus, Haplogroup R-M124 has been observed among Arabs at low frequencies in 11 countries/territories (Egypt, Jordan, Kuwait, Lebanon, Palestine, Qatar, Syria, Tunisia, United Arab Emirates, and Yemen) of the 22 Arab countries/territories so far. In the Kingdom of Saudi Arabia so far has one family identified to have Haplogroup R2A (R-M124) of its paternal genome or Y-Chromosome updated 5 January; 2018.” ref
“In Kazakh tribes it varies from 1% to 12%, however it is found at a higher percent at about 25% among Tore Tribe/ Genghis Khans descendant tribe. In Central Asia, Tajikistan shows Haplogroup R-M124 at 6%, while the other ‘-stan’ states vary around 2%. Bartangis of Tajikistan have a high frequency of R-M124 at about 17%, Ishkashimi at 8%, Khojant at 9%, and Dushanbe at 6%. Specifically, Haplogroup R-M124 has been found in approximately 7.5% (4/53) of recent Iranian emigrants living in Samarkand, 7.1% (7/99) of Pamiris, 6.8% (3/44) of Karakalpaks, 5.1% (4/78) of Tajiks, 5% (2/40) of Dungans in Kyrgyzstan, 3.3% (1/30) of Turkmens, 2.2% (8/366) of Uzbeks, and 1.9% (1/54) of Kazakhs.” ref
“Among regional groups, it is found among West Bengalis (23%), New Delhi Hindus (20%), Punjabis (5%) and Gujaratis (3%). Among tribal groups, Karmalis of West Bengal showed highest at 100% (16/16) followed by Lodhas (43%) to the east, while Bhil of Gujarat in the west were at 18%, Tharus of north showed it at 17%, Chenchu and Pallan of south were at 20% and 14% respectively. Among caste groups, high percentages are shown by Jaunpur Kshatriyas (87%), Kamma Chaudhary (73%), Bihar Yadav (50%), Khandayat (46%)and Kallar (44%).” ref
“It is also significantly high in many Brahmin groups including Punjabi Brahmins (25%), Bengali Brahmins (22%), Konkanastha Brahmins (20%), Chaturvedis (32%), Bhargavas (32%), Kashmiri Pandits (14%) and Lingayat Brahmins (30%). North Indian Muslims have a frequency of 19% (Sunni) and 13% (Shia), while Dawoodi Bohra Muslim in the western state of Gujarat have a frequency of 16%, and Mappila Muslims of South India have a frequency of 5%.” ref
“The R2 haplogroup in the northern regions of Pakistan is found among Burusho people (14%), Pashtuns (10%) and Hazaras (4%). In southern regions, it is found among Balochis (12%), Brahuis (12%) and Sindhi (5%). The R2-M124 haplogroup occurs at a considerably higher rate in the northern regions of Afghanistan (11.4%). Although the true percentage remains debated, the haplogroup is known to be at elevated levels in the Pamiri population (number ranges from 6-17% depending on the group). One study on Nuristanis shows a 20% frequency of R2 (1/5), albeit with a small sample size. 38% of the Sinhalese of Sri Lanka were found to be R2 positive according to a 2003 research.” ref
“While R1a and R1b haplogroups are often associated with the Indo-European languages, the original linguistic affinity of the R2 descendant haplogroups R2a and R2b remains unknown. There are not many studies focusing on deeper subclades of Y‑DNA haplogroup R2 (R-M479), and research tends to focus on the widespread subclades of haplogroup R1. Ancient DNA samples carrying pure Y-DNA haplogroups R1* and R2* haven’t been discovered yet. The oldest detected sample of R* related to both R1 and R2 is the 22000 BCE Mal’ta Boy (MA1), who lived near the Lake Baikal and in archaeogenetics he represents the population of the Ancient North Eurasians (ANE).” ref
“The Y-DNA haplogroup R2 is mainly represented by its R2a (R2‑M124; R‑M124; R‑Y3399; R‑P249; R‑L266) subclade detected in 865 samples worldwide, while the R2b (R‑FGC50368; R‑SK2164; R2b‑FGC21706) subclade was seldom tested for and it was detected in only 13 samples worldwide. In Europe the Y-DNA haplogroup R2 is represented solely by the subclades of R2a (R‑M124) and peaks at 2% in Latvia, 1% in Lithuania, 1% in Belarus, 1% in Masovian Voivodeship of Poland, 1% in Slovakia, 1% in Ukraine, 1% in Moldova, 1% in Hungary and 1% in Switzerland. It does not appear at all in Romania, Serbia, Bulgaria, Scandinavia, Finland, Estonia, Western Poland, Germany, Austria, Ireland, Netherlands, Belgium, France, Spain and Portugal.” ref
R2a in Ancient Samples
- 8241 BC – Ganj Dareh, Iran – Y‑DNA R2a (R‑Y3399), mtDNA unknown (sample I1949)
- 8000 BC – Ganj Dareh, Iran – Y‑DNA R2a (R‑Y3399), mtDNA J1c10 (sample I1945)
- 3500 BC – Geoksyur, Turkmenistan – Y‑DNA R2a (R‑Y8766), mtDNA J1d6 (sample I8526)
- 2196 BC – Gonur, Turkmenistan – Y‑DNA R2a (R‑Y3399), mtDNA R2 (sample I4087)
- 1971 BC – Sappali Tepe, Surxondaryo, Uzbekistan – Y‑DNA R2a-Y3370, mtDNA K1a (sample I7492)
- 1613 BC – Bustan (Boʻston), Uzbekistan – Y‑DNA R2a3a2, mtDNA HV2a2 (sample I11520)
- 1600 BC – Sumbar, Turkmenistan – Y‑DNA R2a-FGC13203 (from R2a‑Y3370), mtDNA W3b (sample I6675)
- 1207 BC – Katelai (Swat), Pakistan – Y‑DNA R2a3a2b2c, mtDNA U7b (sample I12147)
- 400 BC – Saidu Sharif (Swat), Pakistan – Y‑DNA R2a3a2b, mtDNA U2b2 (sample I7722)
- 356 BC – Aligrama (Swat), Pakistan – Y‑DNA R2a3a2b2b1, mtDNA HV6 (sample I8245)
- 166 BC – Ksirov (Danghara Plain), Tajikistan – Y‑DNA R2a (R‑Y3399), mtDNA U2e1e (sample I12292; Kushan)
- 670 AD – Roopkund, Uttarakhand, Northern India – Y‑DNA R2a3a2b2c, mtDNA HV14 (sample I3352)” ref
“The earliest sample containing Y-DNA R2a known to modern archaeogenetics is a man from Ganj Dareh, Iran. This ancient site belongs to the Neolithic Fertile Cresent region, which also contained sites such as Göbekli Tepe, Çatalhöyük, Chogha Golan, Chogha Bonut, Chogha Mish, Abu Hureyra and Jericho. A woman from Ganj Dareh (sample GD13a) shows genetic affinity with Baloch and Brahui from Pakistan and Makrani from Gujarat, India (Baloch mercenaries). R2a carrying male (sample I1945) from Ganj Dareh also carried the descendant haplogroup of mtDNA J1c, which nowadays appears only in Europe and North America, this points to a common origin of his ancestors in a population related to the Ancient North Eurasians.” ref
“Other males from the 3500 BCE Geoksyur site in Turkmenistan carried the following Y-DNA haplogroups: P1 (or CT), 3x J‑ZS6592 (J1a2a1b1a), J‑ZS4428 (from J1a2a1b), J‑PF4993 (from J2a1a1a1a), Q-L56 (Q-M346; Q1a3)[4]. Just like the mtDNA haplogroup J1c, the Y-DNA haplogroup Q1a3 is nowadays widespread mostly among Native Americans, which points to the common ANE origin of Y-DNA P1, R2a, and Q1a3 found at the Central Asian site of Geoksyur. This observation might also explain why in the 2021 Zhang et al. study the Tarim Mummies were modeled as 100% isolate but their genes were also found not only in the Neolithic Central Asian Geoksyur, Namazga, and Sarazm individuals but also in the Botai, Okunevo, Dzungaria, Chemurchek, Bronze Age Baikal and Bronze Age Kumsay in Kazakhstan. They all shared a high level of Ancient North Eurasian (ANE) ancestry.” ref
“Nevertheless, the samples from Geoksyur, Namazga, Parkhai, and Sarazm were still much closer (but not uniform) to Yamna Samara and Afanasievo Culture than to Tarim Mummies and Mal’ta Boy. The only samples from Central Asia shifted towards ANE the most were those from Bronze Age Aigyrzhal, Kyrgyzstan (circa 2100 BCE) and resembled the ones from Early Bronze Age Dzungaria. The downward shift of Geoksyur and Namazga from Yamna Samara was mostly due to its high genetic input from Iran Neolithic and Anatolia Neolithic, in evidence from that earliest 8211 BCE Ganj Dareh sample and lots of Y‑DNA J1a and J2a in Southern Central Asia. That is why the pre-Steppe carries of Y‑DNA R2a in Central Asia can be modeled in aDNA as 10% ANE, 20% Anatolia Neolithic, and 70% Iran Neolithic (that is 90% of Fertile Crescent or Zagrosian DNA). The Elamites might be genetically closest to people from the Neolithic Central Asia but this requires some further examination.” ref
“The phenetic analysis of Tarim Mummies from Bronze Age Qäwrighul (1800 BCE) also pointed to their uniqueness and to their slight affinity towards people from the Oxus Civilization (BMAC) and Sappali Tepe. However, no Y‑DNA R2a has ever been detected in any of the Tarim Mummies from the Xiaohe Cemetery (2100 BCE – 1700 BCE), and instead, they carried mainly the descendants of Y‑DNA haplogroup R1*: 2x R1b‑PH155 (R1b2), 7x R1a1a, 1x R1‑M173. The remaining not deeply classified Y‑DNA haplogroups from Xiaohe were: F‑M89 (East, South and Central Asian), K‑M9 (ancestral to N, O, P, R; 43000 BCE Ust’-Ishim man carried a descendant of this haplogroup: K2a), P‑M45 (ancestral to Q and R).” ref
“BMAC individuals were largely derived from local, Chalcolithic populations similar to those found in Geoksyur. A genome of a woman from 2800 BCE – 2300 BCE Rakhigarhi, Northern India (close to Punjab and New Delhi) matched the DNA from 11 other individuals who had been found at sites in Iran and Turkmenistan from a similar period. A comparison of modern Indo-Iranian populations with the ancient genome has shown genetic continuity with the BMAC cluster, except for their high admixture with Steppe populations (Andronovo Culture), that occurred at the end of BMAC after 2000 BCE. The first genetic outliers observed in several BMAC individuals may represent the very beginning of this demographic event. According to Narasimhan et al. 2019, Steppe pastoralist ancestry appeared in outlier individuals at BMAC sites by the turn of the second millennium BCE around the same time as it appeared on the Southern Steppe (in Sintashta Culture and Andronovo Culture).” ref
“The Central Asian Steppe group, which admixed with BMAC does not harbour the Baikal Hunter-Gatherer genetic component because the BHG component did not arrive in the Central Asian Steppe until the late Iron Age. The best proxy for admixture with BMAC is the subpopulation formed by four individuals all belonging to the Kytmanovo site of Andronovo Culture (1446 BCE – 1298 BCE male from Kytmanovo already carried Y‑DNA R1a‑Z2124). These individuals have a very similar genetic profile to that of individuals from Sintashta, near the Caspian Sea. The screening of the individuals associated with the Andronovo horizon in the data shows that they formed a moderately heterogeneous group, with genetic proximities that do not necessarily correlate with geographical or cultural proximity. For example, Srubnaya-Alakulskaya individuals are more closely related to individuals grouped under the precise Andronovo label than Srubnaya from the Samara region.” ref
“The 2200 BCE – 1600 BCE males from Sappali Tepe, Gonur, Bustan (Boʻston), and Sumbar, who carried the Y‑DNA haplogroup R2a still archaeologically belonged to the BMAC (Oxus Civilization). In the post-1200 BC Swat Valley, Pakistan males carrying R2a appear next to males, who carry the Sintashta derived R1a‑Z94 haplogroup (from R1a‑Z93). None of the Yamna/Afanasievo derived Y‑DNA R1b‑Z2103 appeared in the Swat Valley, even if some males from Sintashta still carried this Y‑DNA haplogroup (2012 BC sample I1020[4]). North of Pakistan there are three additional ancient samples with detected Y‑DNA R1a‑Z93: one from 1497 BCE Kokcha, Uzbekistan (sample I12499) and two from 1200 BCE Kashkarchi, Uzbekistan. One of males from Kokcha, Uzbekistan from around 1738 BCE carried Y‑DNA haplogroup Q1a1b1, which is related to both Native Americans, and Siberian Kets, yet another sign of some Ancient North Eurasian derived genes in Central Asia.” ref
“A 1207 BCE man from Katelai (Swat), Pakistan carried both Y‑DNA R2a3a2b2c and mtDNA U7b. The modern distribution of mtDNA haplogroup U7 almost perfectly matches the distribution of Y‑DNA haplogroup R2 shown on the map above (and below). This supports the hypothesis of a local origin of Y‑DNA R2a and R2b in the Swat Valley of Pakistan because the carriers of mtDNA U7 spread to South Asia and Europe before the suggested Bronze Age expansion of Indo-European languages from the Andronovo Culture and mtDNA U7 originated in the Near East and then spread towards South and Central Asia prior to the Holocene. However, low amounts of mtDNA U7 in West Bengal and Orisa most probably deny the appearance of mtDNA U7 together with Y-DNA R2a in South Asia (but not in Central Asia or Pakistan), and it is still more likely that Y-DNA R2a spread to East India together with Y‑DNA R1a‑Z93, which came from the Andronovo Culture and then those two Y‑DNA haplogroups spread together from the Swat Valley of Pakistan. The latter is also supported by the elevated amounts of mtDNA U7 in Gujarat and Rajasthan and much lower amounts of Y‑DNA R2a there.” ref
“Ancient individuals from the Swat Valley of Pakistan show that the Andronovo Culture (Central Asian Steppe) ancestry integrated further South around 1500 BCE – 1200 BCE, and nowadays this genetic component makes up to 30% of ancestry in many South Asian populations. That Central Asian Steppe ancestry in South Asia has the same genetic profile as that of the Bronze Age Eastern Europe, tracking a movement of people, who affected both regions and likely spread the unique features shared between Indo-Iranian and Balto-Slavic languages.” ref
R2b in Ancient Samples
- 4000 BCE – Anau Tepe (Tepe-Anau), Turkmenistan – Y‑DNA R2b (R‑FGC50368), mtDNA W3a1 (sample I4087)
- 850 BCE – Loebanr (Swat), Pakistan – Y‑DNA R2b (R‑FGC50368), mtDNA W3a1b (sample I8997)
- 850 BCE – Loebanr (Swat), Pakistan – Y‑DNA R2b (R‑FGC50368), mtDNA W3a1b (sample I8998)” ref
“Those two males from Loebanr were most probably brothers from the same mother, and again, the oldest subclades of Y‑DNA R2 appear near the Swat Valley and in Central Asia instead of South India (or South Asia in general). The oldest sample of mtDNA W3a1 in Central Asia comes from 4000 BCE or around 6,000 years ago, a male from Anau Tepe, who also carried Y‑DNA haplogroup R2b just like those two brothers.” ref
“The second oldest sample carrying mtDNA W3a1 comes from a 2917 BCE Yamna Culture woman from Prydnistryanske, Ukraine (very close to Moldova). Bronze Age samples of that mtDNA haplogroup come mostly from European and Central Asian cultures such as Corded Ware, Baden, Unetice, Trzciniec, BMAC (Sappali Tepe, Uzbekistan), Scythians from Kazakhstan. Its sister haplogroup W3a2 is nowadays found mostly in Tajikistan, England, France, Slovakia, Romania, Poland and Ukraine.” ref
“The 2022 paper by Guarino-Vignon et al. showed that the Neolithic and Chalcolithic samples from Central Asia, including those from Tepe-Anau genetically resemble modern Brahui, Balochi, Makrani, Kalash and Pathans (Pashtuns) but modern Yaghnobis and Tajiks are dramatically shifted from those samples towards Yamna and Iron Age Turkmenistan (Scythians, Kushans). Some samples from Early Middle Bronze Age Turan (circa 1500 BCE) resemble those from Central Asian Steppe (Andronovo Culture) and those of modern Ossetians. The Burusho people appear closer to Pathans and Kalash but form their own cluster far away from the ancient Central Asian samples and Tajiks. The Burusho are still closest to the South Asian cluster.” ref
“Modern samples of R2b were only detected in Pakistan (Islamabad; Punjab; Azad Kashmir), India (Punjab), China (Shandong), Iraq (Al Anbar), Saudi Arabia (Hāʼil), Bahrain (Al Muharraq). It is possible that this haplogroup appeared in Saudi Arabia, Iraq and Bahrain already in the Neolithic but the Iron Age Persian expansion or some other recent events could also be responsible for spreading R2b to those regions. This can not be determined.” ref
R2a in Kazakhstan and Europe
“The relatively high amount (0% – 25%) of Y‑DNA R2a in the Kazakh Tore tribe, 12% in Zhanakorgan district, 6.3% – 10% in Kozha tribe, 5.4% in Jalayir tribe, 2% in Sirgeli and Alban tribes, 1% in Baiuly tribe could be explained by the backward migration of Scythians from the territory of BMAC and Southern Andronovo (there are no ancient samples of Y‑DNA R2a from the territory of Kazakhstan). Yaghnobis (cultural and genetic descendants of Scythians) show no genetic admixture from South Asia (unlike Tajiks), while both Yaghnobis and Tajiks show genetic affinity to BMAC and Andronovo[10]. This backward Scythian migration through Kazakhstan is further proven by 8% of Y‑DNA R2a in Ossetians from Alagir, 7.9% in Balkars (descendants of Scythian Alans and Turkic tribes; they carry very high amounts of Central Asian Y‑DNA R1b‑M73 and Iranic Y-DNA R1a‑Z2124 from R1a‑Z94), 16% in Chechens, 20% in Kalmyk-Oirats from Derbet, Kalmykia and 1% in Eastern European countries.” ref
“During the 14th century CE, Alania was destroyed by Timur, and many of the Alans, Cumans, and Kipchaks migrated to Eastern Europe. This genetic influx might go back even further in time as R1a‑Z2124 was already detected in the ancient Hun (5th century CE), Avar, and Hungarian conqueror (post-850 CE) samples from Hungary, a descendant of this Y‑DNA haplogroup, namely the R1a‑Z2123 is nowadays mostly prevelant in Bashkiria, Kazakhstan (in Tore, Kozha, Tarakty, Sirgeli, Oshakty, Kangly, Jalayir, Zhetiru, and Kypshak tribes), Northern Afghanistan, Tajikistan, among Karachay-Balkars and it was the Y‑DNA haplogroup of the Hungarian Árpád Dynasty.” ref
“Even older sample with this Y‑DNA haplogroup is a 380 BCE – 200 BCE Scythian from a kurgan near Samara (I0247), who carried Y-DNA R1a1a1b2a2a (R1a‑Z2125) and mtDNA G2a4. Both Y‑DNA haplogroups R1a‑Z2124 and R1a‑Z2125 were present in the territory of the Middle Bronze Age Kazakhstan because a 1900 BCE – 1300 BCE male from Dali in Bayan-Zhurek mountains, Eastern Kazakhstan carried Y‑DNA R1a‑Z2124 and mtDNA U5a1a2a (sample I0507), this site is only 100 km away from Northern Xinjiang. Another male from 1600 BCE – 1196 BCE Taldysay, Ulytau region, Central Kazakhstan carried R1a‑Z2125 and mtDNA H3g (sample I4787).” ref
“The Kalmyk Dörwöds (Dörbet) in Mongolia are characterized by high frequencies of Y‑DNA haplogroups C3d‑M407 and R2a‑M124 (15.2%). It is noteworthy that the age of haplogroup R2aM1‑24 found in both Dörbet and Oirats from Kalmykia shows the best correspondence to the historical chronology of the Kalmyk dispersals. Haplogroup D-M174, albeit rarely found in the Dörwöds and Torguuds (about 2%), reveals genetic affinity to the Tibetan populations, where this haplogroup was present probably before the Last Glacial Age. Another rare Y‑DNA haplogroup N1a‑M128, which frequently occurred only among Kazakhs (8.1%) in Central Asia, was also detected in the Buzava Kalmyks, who live next to Derbet in Kalmykia and carry up to 17.4% of Y‑DNA haplogroup R2a.” ref
“Phylogenetic network of haplogroup R1a1a‑M17 (carried by 3.3% of Kalmyks) demonstrates that two haplotypes of the Kalmyks coincide with the haplotypes common among different populations of South Siberia (Tuvinians, Altaians and Sojots) and Central Asia (Tajiks and Pathans), two haplotypes with populations of Pathans, Persians and Indians, and one haplotype with South Siberians (Shors and Tuvinians).” ref
“A median network of R2a‑M124 haplotypes derived from 12 Y‑STR loci in the Kalmyks, Buryats (2.7% of R2a) and populations of Southwestern and South Asia (in which this haplogroup is present at a relatively high frequency) reveals common haplotypes observed in Kalmyks and Buryats, whereas there is no haplotype sharing between these and other comparative populations. However, the R2a‑M124 haplotypes found in the Kalmyks and Buryats are still the derivatives of Indian subclades of R2a. This suggests a recent common ancestry and/or expansion of the Kalmyk R2a‑M124 lineages from India or Southern Central Asia.” ref
“The descendants of Y‑DNA R2a‑F2791 subclade are nowadays found only in Latvia, Ukraine, Slovakia, Chechnya, Georgia, Kuwait, Saudi Arabia, India, Pakistan, Afghanistan, and Iran. The R2a‑Y101424 subclade of R2a‑Y3399 (basal R2a‑M124) found among Kurds in Turkey is also present in Iraq, Pakistan, Lebanon, Uzbekistan, and Saudi Arabia, so it most probably was not mediated there by the Scythians. Kurmanji Kurds from Georgia carry around 44% of Y‑DNA R2a‑M124. The Y‑DNA haplogroup R2a has not been detected in the Roma population of Europe[35]. They mostly carry: 44.8% of H‑M82, 22.6% of I‑M170, 12.7% of J2a‑M67, 6.7% of R1‑M173, 3.6% of E‑M36, 2% of J‑M92, 1.6% of C‑M217, 0.4% of R‑M17 (R1a1a) and aren’t responsible for the appearance of Y‑DNA R2a in Europe (the Scythians are and from the 17th century CE the Kalmyks).” ref
R2a in China
“Archaeological excavations at the 2000 BCE site of Sappali Tepe, located in the North Bactrian oasis of Southern Uzbekistan, during the 1960s and 1970s yielded the earliest evidence of silk outside of China and raised the possibility that the East‑West contacts along the Great Silk Road may be far older than previously thought. People from the Southern Gansu province of China nowadays carry 3.4% of Y‑DNA haplogroup R2a. During the Bronze Age, the Qijia Culture (2200 BCE – 1600 BCE) thrived in that region, and it might be one of the factors why the R2a haplogroup spread to Central China.” ref
“Qijia Culture is distinguished by the presence of numerous domesticated horses (in evidence also genetically after DOM2 horse spread to China after ca. 2000 BCE), practice of oracle divination, metal knives, and recovered axes, which point to some interactions with Siberian and Central Asian cultures, in particular with the Seima-Turbino complex (Okunevo Culture). Archaeological evidence points to plausible early contact (often violent) between the Qijia Culture and Central Asia. In spite of all the archaeological evidence of its Western contacts, those most probably were only trade relations as all males from the 2000 BCE Mogou site of the Qijia Culture carried only East Asian Y‑DNA haplogroups: O3a2 and O3a2c1a and did not show any genetic affinity towards West Eurasia. It means that Y-DNA haplogroup R2a appeared in Central China only later in history. 4% of R2a in Western Xinjiang and 2% in Eastern Xinjiang might point to its appearance in that region after 1500 – 1300 BCE because a study from 2021 by Ning et al. proved that the appearance of European mtDNA in 1500 – 1300 BCE Western Xinjiang is correlated with the appearance of Yamna (Steppe) and Andronovo related ancestry.” ref
“In the same study, mtDNA samples from the Qijia Culture clustered with East Asian and South Asian samples far away from European samples, this again suggests that the speculated appearance of Y‑DNA haplogroup R2a in China during the Early Bronze Age either wasn’t correlated with the appearance of Yamna or Andronovo related ancestry or did not happen during the Qijia period in Gansu. The Qijia mtDNA samples were not far away from the Late Bronze Age Kayue, China; Bronze Age Khovsgol, Mongolia and Neolithic samples from Baikal, Russia. Those three geographic regions nowadays harbor around 3% of R2a. Kayue Culture (900 – 600 BCE) most probably developed from the Western part of the Qijia Culture. Early Bronze Age Tarim (Xinjiang) clustered with the Kayue Culture and Neolithic Baikal but not with the European samples.” ref
“The only Asian ancient mtDNA samples clustering close to those from Europe were: Eneolithic Turkmen (Namazga Culture), BMAC, MLBA Central Asian Steppe, Iron Age Shirenzigou and Northern Xinjiang Afanasievo (a proof that Tocharians and Yuezhi are the descendants of the Afanasievo Culture). After 1000 BCE the stone based burial type spreads from Northern Xinjiang Dzungaria (Afanasievo mtDNA) to Eastern Xinjiang, this might represent the earlieast spread of Tocharian languages to Eastern Xinjiang noted by archaeology. According to phenetic comparisons, the Tarim Basin (Xinjiang area) experienced a significant gene flow from highland populations of the Pamirs and Fergana Valley (modern-day Eastern Uzbekistan) after 1200 BCE.” ref
“These highland populations may include those who later became known as the Saka (Scythians) and who may have served as “middlemen” facilitating contacts between the East (China) and West (Bactria) along what later became known as the Great Silk Road. The ancient Scythian DOM2 horse belonged to exactly the same breed as that of Boz Adyr, Xiongnu (from Gol Mod 2 Cemetery, Mongolia), and Karasuk, all those breeds were also closely related to the DOM2 horse from Sintashta, meaning that the people responsible for spreading the final breed of DOM2 horse to China were the Scythians. This lineage of horses survived at least until the 8th century CE in Central Asia at Boz Adyr, Kyrgyzstan. Bronze Age Deer Stone horses from Mongolia, medieval Aukštaičiai horses from Lithuania (9th – 10th century CE), and Iron Age Pazyryk Scythian (6th century BCE) horses showed similar diversity levels.” ref
“This might point to the earliest possible date of 1200 – 700 BCE for the arrival of Y-DNA R2a in the Tarim Basin and points to a similar backward migration of Scythians to the territory of late Andronovo Culture, which also took place in Kazakhstan. The Saka (Scythian) presence has been found in various locations in the Tarim Basin, for example, in the Keriya region at Yumulak Kum (Djoumboulak Koum, Yuansha) around 200 km East of Khotan, with a tomb dated to as early as the 7th century BCE. Surviving documents from Khotan (300 BCE – 1006 CE) of later centuries indicate that the people of Khotan spoke the Saka language (Khotanese), an Eastern Iranian language that was closely related to the Sogdian language from Central Asia. Additionally, the mtDNA samples from Iron Age Western Xinjiang and Southern Xinjiang were shifted from the Bronze Age Xinjiang samples towards BMAC, Tian Shan Saka, Tagar (Scythians), Iron Age Sarmatian, and Hungary Bronze Age.” ref
“According to the 2022 V. Kumar et al. study on ancient DNA from Xinjiang (201 samples), the Bronze Age (3000 – 1000 BCE) Northern and Western Tarim Basin is mainly represented by Y-DNA haplogroups: R1b1 (majority), R1a2 (it is either very rare R1a‑YP4141 or R1a‑Z93), Q2a, Q2, and Q1b1. The Late Bronze Age in the Western Tarim Basin (probably circa 1000 BCE) yields the first appearance of Y‑DNA R1a1. It is during the Iron Age (1000- 1 BCE) when the majority of Y‑DNA haplogroups related to Central Asia appear in Western Tarim Basin: R1a1 (majority), R1b2 (R1b‑PH155), Q2a1, Q1b2, Q1b1, J2a1, J2a2, J2b2, J1a2, L1a2, E1b1 (E1b1b1 was already found at the 2011 BCE Gonur site of BMAC, so it is not Greek or Macedonian), I2a1 (European). The Iron Age Southern Tarim Basin is dominated by Y‑DNA: G2a2 (majority), R1a1, L1a1, Q1b1 (majority), Q1b, Q1, N1a1, and Iron Age Northern Tarim Basin by Y‑DNA: R1b1, R1a1, and R1a2.” ref
“This means that this study, in opposition to 2021 Zhang et al., proves that the Tocharian languages (R1b1) were first in the Tarim Basin right next to some isolated unknown ANE languages (Q2a, Q1b), and exactly after 1000 BCE or around 3,000 years ago, the Scythian genetic influx occurred in Western Xinjiang. The Y‑DNA haplogroup O2 appears in the Eastern Tarim Basin only during the Iron Age and in the Southern Tarim Basin much later in the historical era (1 CE—present). Deeper classification of some of those detected Y-chromosomes on YFull showed that two Xinjiang individuals from 1451 – 1374 BCE and 739 – 494 BCE already carried the Y‑DNA haplogroup R1a‑Z93; two from 1571 – 1460 BCE and 353 ‑ 7 BCE carried the Iranic Y‑DNA R1a‑Z2123. The R1b‑FT292014 haplogroup from Xinjiang is nowadays found only in Pakistan, while R1a‑YP1548 from a 693 BCE Xinjiang man is found only in Kazakhstan, Altai, Kyrgyzstan and Shandong, China (next to 6.7% of R2a in Jiangsu, China).” ref
“Surprisingly, on aDNA PCA, four Late Bronze Age and two Bronze Age Tarim Basin individuals were identical to modern Europeans. Remaining BA samples from Tarim Basin clustered with Early Bronze Age Dzungaria (Proto-Tocharians). Those people might literally be the ancestors of Tocharians, and those 6 people “from Europe” were either European mercenaries/merchants or individuals who still preserved their Yamna/Afanasievo gene pool (maybe they did not mix with the ANE-derived locals). Samples of EMBA (1700 BCE) Tarim Mummies from the 2021 Zhang et al. paper created their own cluster far away from the Afanasievo/European Tarim Basin population and appeared closer to the supposed ancestral ANE population. During the Iron Age, the Tarim Basin samples shifted towards Iran Neolithic (BMAC) and East Asia but still did not move to the exact area of modern Uyghur, Iron Age Southern Tarim Basin, and Iron Age Shirenzigou samples.” ref
“When it comes to Horpa (Qiangic) speaking population of Daofu, Western Sichuan, China, where R2a makes up 6.25% of all Y‑DNA, on PCA they appear to be closest genetically to Han-Xinjiang and Tibetan-Shigatse populations. The European mtDNA haplogroups were not found among Horpa (Qiangic) speaking populations. The mtDNA (maternal) variation of Qiangic populations was also largely contributed by Northern Asian prevalent haplogroups, including haplogroups A, C, D, and G. In addition, cultural features of the upper Yellow River basin, such as painted pottery, millet agriculture, and urn burial, are prevalent in the Neolithic sites of Western Sichuan, probably due to the demic diffusion via the genetic corridor.” ref
“Hui from Ningxia (close to Gansu and Henan) carry 3.2% of R2a, they also carry 1.6% of R1b1b2 (R1b‑M269) and 3.2% of R1a1, unlike Man from Liaoning, who carry 1.5% of R2a but none of the Y-DNA R1 derived haplogroups (just like Qiang from Sichuan and Han from Gansu who carry only R2a). Han from Jiangsu also carry 8.9% of R1a1 next to 6.7% of R2a. In general, R1a and R1b are not totally absent in China as Hui from Yunnan carry 10% of R1b1c and 10% of R1a (next to 10% of J1, 10% of Q1a3, and 0% of R2a). Chinese from the Henan province (next to Gansu) also carry around 1.6% of R1b‑M269.” ref
“R1b1b1 (R1b‑M73 brotherly to R1b‑M269) has also been detected in 8.3% of Chinese Mongolian, in 25% of Kyrgyz from Xinjiang, and in 6% of Uyghurs from Xinjiang (they also carry 30% of R1a1, 16% of J2a and 4% of R2a). R1b‑M73 was detected mainly in NEAS (Kumandins, Mongolians, Chinese, Bashkirs, Northern Pakistani, and Pamiri) and was sporadically detected in South Asia and West Eurasia (except for Karachay-Balkars and Kalmyks). It is possible that this haplogroup, together with R2a and R1a‑Z93, spread to those regions with migrations of the Saka after 1000- 700 BCE and later with those of Xiongnu, Wusun, Yuezhi (probably to Gansu) or Kushans after the 3rd century BCE. In general, the star-like structure of Y‑DNA R2a‑M124, which includes the Chinese samples, suggests that it reflects a relatively recent expansion from South or Central Asia to North-East Asia via the Northern route and more diversified R2 haplotypes occur in the areas around Pakistan and India instead of North-East Asia.” ref
R2a in Thailand and Malaysia
“The Mon from Northern Thailand shows the presence of haplogroups usually found in South, Central, and West Asia: 16.7% of R‑P249 (R2‑M124) and 5.6% of J‑M172 (J2). The connection between the ethnic Mon and populations from South and Central Asia was already proposed from previous identification of mtDNA lineage W3a1b. Both groups of the Mon-Khmer speaking Lawa groups showed high differences between each other and from other populations, presenting low levels of haplogroup diversity with high frequencies of O‑PK4 (O1b1a1) (72% in Lawa 1) and N‑M231 (56% in Lawa 2).” ref
“The native language of Mon is Mon, it belongs to the Monic branch of the Mon-Khmer language family and shares a common origin with the Nyah Kur language, which is spoken by the people of the same name and who live in the Northeastern Thailand. The groups from Thailand, where Y‑DNA haplogroup R1a has been detected are: Khon Muang 6 (9% of R1a; 4.6% of J2; 0% of R2a), Yuan 1 (5.3% of R1a; 0% of J2; 0% of R2a), Yuan 3 (4.2% of R1a; 0% of J2; 0% of R2a). It means that the appearance of R2a in Thailand could have been accompanied by the spread of Y‑DNA haplogroups R1a (usually R1a‑Z94 derived R1a1a1b2a1b: R1a‑Z2124) and J2.” ref
“Y‑DNA R2a (R‑L266, same as R‑M124) and mtDNA R21 has been detected in one modern male (JEH002) from the Malaysian Jehai tribe, who live next to the Thailand’s Southern border. Other male (JEH04) from the same tribe carried Y‑DNA R1a1a1b2a R1a‑Z94 and mtDNA M13b1. If the appearance of Y‑DNA R2a correlates with that of R1a‑Z94 in Thailand and Malaysia, then the spread of R2a in Thailand could occur only after 1200 BCE (after R1a‑Z93 associated Painted Grey Ware Culture appeared in Central India next to West Bengal) but this can not be determined yet. Ancient DNA samples from Thailand carrying Y‑DNA R2a could undermine this statement but just like in the case of ancient samples from Southern India, there aren’t any carrying Y‑DNA R2a.” ref
Languages and Genetics
“Modern distribution of Y‑DNA haplogroups R2a and R2b covers the areas of not only the Indo-European languages but also: Dravidian Brahui in Pakistan; isolate Burushaski in Hunza, Pakistan; Horpa (Qiangic) near Xinlong, Western Sichuan, China; Monguor in Gansu, China; Mandarin in Jiangsu, China; Mon in Thailand; Arabic in Jordan, Kuwait and United Arab Emirates; Jehai (Mon-Khmer) in Malaysia; Kalmyk-Oirat, Buryat (Mongolic), Karachay-Balkar, Ossetian and Chechen in Russia; Kazakh in Kazakhstan; Oirat in Western Mongolia; Lodha, Kannada, Kerala, Malayalam, Tamil, Telugu and Sora in India.” ref
When it comes to Indo-European languages the distribution of Y‑DNA R2 covers the areas of: Tajik, Bartangi and Wakhi (Pamir languages) in Tajikistan; Nuristani, Pashai, Hazara and Aimak in Afghanistan; Balochi, Pashto, Urdu and Punjabi in Pakistan; Persian and Balochi in Iran; Kurdish in Turkey, Iraq, Iran; Kurmanji in Georgia; Dhivehi in Maldives; Sinhalese in Sri Lanka; Kashmiri, Hindi, Bhojpuri, Bengali, Odia, Punjabi, Marwari, Marathi (in Konkan) and Konkani languages in India. All those languages belong to the the Indo-Iranian branch of the Indo-European languages.” ref
“R2a is only weakly related to the broad distribution of the Hindi language and in areas of Dravidian languages its amounts reach from 3% to 10%. Even if this Y‑DNA haplogroup predates the Yamna (steppe) related genetic input in Central Asia it can not be associated with the appearance of Indo-European languages in Western North India, especially in Gujarat, Rajasthan and Madhya Pradesh.” ref
“Languages mentioned above from the areas where R2a exceeds 14% are marked in bold. From those the ones closest geographically to the appearance of Y‑DNA R2 in ancient samples are: Pamir, Punjabi, Burushaski and Nuristani languages. Václav Blažek in his 2019 paper “Toward the question of Yeniseian homeland in perspective of toponymy” suggests that the Pamir languages have a Burushaski-like substratum. Although Burushaski is today spoken in Pakistan to the South of the Pamir language area, Burushaski formerly had a much wider geographic distribution before being assimilated by Indo-Iranian languages.” ref
“In 203 Burushaski words I found only 15 (that is 7.4%), which might be related to the Indo-European languages: pʰu (“fire”), sua (“good”), ga’rurum (“hot”), mi (“we”), ja (“I”), te (“that”), hir (“man”; Latin: vir), mosh (“nose”), ren (“hand”), girashom (“play”; Old Church Slavonic: igrati; Lower Sorbian: graś), jee (“live”), sa (“sun”), thas (“smoke”), tik (“earth”; Hittite: tekan; Tocharian A: tkaṃ). Similar amount of words in common appears between the Indo-European and Native American languages associated with Y‑DNA haplogroup Q (brother of R* from ANE; 5/96 in the words section of my website resulting in only 5.2% of shared vocabulary).” ref
“It means that the Burushaski language is still a valid candidate for the language most likely associated with the Y‑DNA haplogroup R2. Other most common Y‑DNA haplogroup among Burusho is R1a1 (26%) and from those 15 “Indo-European” Burushaski words only 4 are not in common with Sanskrit. Burushaski words not shared with Sanskrit but instead shared only with the European IE languages are marked in italic bold, and they make up 2% of vocabulary shared between the Y‑DNA R1b‑M269/R1a‑Z283 associated languages and Y‑DNA R2a “associated” Burushaski.” ref
“Other most frequent Y‑DNA haplogroups among Burusho are: 12.4% of L‑M357, 4% of L‑M20, 7.2% of J2‑M172, 4% of H, 2% of Q‑M242 (from ANE). Y‑DNA haplogroup L reaching 16.4% among Burusho is the second most likely candidate to be associated with the origin of the Burushaski language. There are also some words in common between Indo-European (R1a and R1b), Horpa (6.25% of R2a) and Khroskyabs / Tangut (from the same language family branch as Horpa), which could possibly go back to the times of the basal R* haplogroup (ANE):
- Horpa (Rgyalrongic): zjar (“heart”)
- Khroskyabs (Rgyalrongic): sjar
- Swedish: hjärta
- Bulgarian: сърце (sǎrce)
- Udmurt: сюлэм (sjulem)
- Horpa (Rgyalrongic): rɛku (“elbow”)
- Lithuanian: ranka (“hand, arm”)
- Latvian: roka (“hand, arm”)
- Polish: ręka (“hand, arm”)
- Burushaski: ren (“hand”)
- Serbo-Croatian: ruka (“hand, arm”)
- Quechua (Y-DNA Q): ruk’ana (“finger”)
- Khroskyabs (Rgyalrongic): stî (“to exist”)
- Hittite: asanta-
- Sanskrit: अस्ति (asti)
- Tangut (Rgyalrongic): khu (“dog”)
- Tocharian A: ku
- Ancient Greek: κύων (kúōn)
- Old Irish: cú
- Hittite: kuwaš” ref
“Again, those similarities do not exceed 2% of shared vocabulary. The languages, which would exceed this barrier are the Indo-Iranian languages but the sole association of those language with only the Y‑DNA haplogroup R2 would result in the rejection of their association with Y‑DNA haplogroup R1a‑Z93. R1a‑Z93 is the only haplogroup, which strongly binds the Indo-Iranian languages with Europe. To say that Sanskrit originated already with haplogroup’s R2 split from R* (circa 14000 BCE) requires an explanation why the Y‑DNA haplogroup R2a is not as common as R1a‑M780 (from R1a‑Z93 but not from R1a‑Z94 or R1a‑Z2125) in Western North India. The Indo-European areas of India with 0% of R2a are Rajasthan and Assam, while in the Punjab disctrict without the Brahmins it reaches only 4%.” ref
“People from Malana in Himachal Pradesh, India speak the Sino-Tibetan Kanashi language and do not carry any Y‑DNA R2a at all but instead they carry: 60% of J2a1h, 27% of R1a, 10% of H, and 3% of L. This high amount of R1a can not be ignored as “the Kanshi seems to be a mixture of Sanskrit and several Tibetan dialects.” ref
“It is important to notice that the distribution pattern of R1a‑Z93 derived haplogroups in South Asia (India, Nepal, Sri Lanka, Maldives) usually matches the distribution pattern of R2a‑M124, for example in Sri Lanka the amounts of R2a and R1a are almost equal, in Uttarakhand the Tharu carry 22% of R1a and 13% of R2a, the Goswami carry 6.5% of R1a and 6.5% of R2a, in Uttar Pradesh there is 19% of R2a and 26.5% of R1a1a. There are some exceptions tough like in Maldives where R1a is more frequent than R2a and in Gujarat where R2a is only 3% – 18% and R1a makes up the majority of Y‑DNA haplogroups there (from 10% to 60% depending on place of measurement)[56], the Brahmins from Uttarakhand carry 46% of R1a and 11% of R2a.” ref
“To say that Iranian languages have nothing to do with R1a‑Z2125 (from R1a‑Z94 and R1a‑Z93) is to ignore the levels of that Y‑DNA haplogroup reaching more than 50% in Tajikistan, Kyrgyzstan (former Saka territory) and Northern Afghanistan (62% of R1a1a, 0.5% of R1b‑L23 and 11% of R2a). The only areas with lower frequencies of R1a‑Z93 are those with higher frequencies of R2a, so there is still some possibility that the Indo-European Nuristani or Pamir languages could have some association with the Y‑DNA haplogroup R2a but this is only a speculation (maybe better explained by their Burushaski substratum). The area of Dardic languages during the Bronze Age (1200 – 500 BCE) in the Swat Valley of Pakistan contained more samples of Y‑DNA haplogroup R2a than what can be found in the people who live in this area today, namely the Kho (speakers of Khowar), Pathan (nowadays 0% of R2a; 50% of R1a) and Kalash (nowadays 0% of R2a; 20.5% of R1a). The aforementioned people from the Swat Valley fit into their own genetically (aDNA) isolated cluster, while the Burusho are close to Kho and Kalash but still form a slightly different cluster. This might be easily explained by much higher amounts of Andronovo derived ancestry in Kho and Kalash.” ref
“A study from 2015 by Ayub et al. showed that the Kalash people are genetically isolated from the South Asian populations and mostly resemble the Ancient North Eurasian Mal’ta Buret Boy (Paleolithic Siberian Hunter-Gatherers). It means that the ancestors of the Kalash arrived in the territory of Northern Pakistan already around 10,000 – 8,600 BCE or around 12,000 to 10,600 years ago. Those ancestors most probably carried the Y‑DNA haplogroups R2 and Q. This does not mean that they did not receive any genetic input after 8600 BCE. Kalash are even more genetically isolated than the Burusho people but still speak an Indo-Iranian language and carry 20.5% of R1a and 6.8% of R*. According to a 2016 study by Lazaridis et al. the Kalash from Pakistan are inferred to have around 50% of Yamna related ancestry, with the rest being Iranian Neolithic, Onge, and Han. The Kalash have a high proportion of Y‑DNA haplogroup L3a lineages, which are characterized by having the derived allele for the PK3 Y‑SNP and are not found elsewhere. They also have predominantly Western Eurasian mitochondrial lineages and no genetic affiliation with East Asians.” ref
“The Khowar language (and with it other Dardic languages, including Chitral Kalasha) can surely be excluded from the proposed cultural descendants of Y‑NA haplogroup R2a because even if it is geographically close to the Burusho, who carry 14.4% of R2a, the Kho and Kalash carry none and the Khowar language still retains many words in forms closest to Sanskrit. The Western Eurasian mtDNA haplogroups (HV8, H19, H57, H24, C, and C4a) were predominantly observed in the Kho samples with an overall frequency of 50%.” ref
“On aDNA PCA, the Kho people are slightly shifted from Burusho and Kalash towards Iranian, Central Asian, Siberian, and European samples. That is why “Kho can trace a large proportion of their ancestry to the population who migrated South from the Southern Siberian steppes (Andronovo Culture) during the second millennium BCE (~110 generations ago). An additional wave of gene flow from a population carrying East Asian ancestry was also identified in the Kho (also in Balti and Burusho) that occurred ~60 generations ago and may possibly be linked to the expansion of the Tibetan Empire during 7th – 9th century CE”. The Kho possess genetic ancestry components associated with the Yamna Culture and European Neolithic Farmers (Anatolia Neolithic + WHG); these three components taken together are characteristic of Middle Bronze Age populations from the Andronovo Culture, while the Early Bronze Age Central Asian Steppe populations lack the additional Anatolian Neolithic and WHG components. The shift of Kho from the 3500 BCE Geoksyur, Turkmenistan samples is also evident.” ref
“The only remaining Indo-European candidates closest to the archaeological sites, where oldest R2a samples were found, would then be the Nuristani languages (some Nuristani males still carry Y‑DNA R2a). To prove that the Nuristani languages in Afghanistan are the descendants of a language spoken by the people who carried the R2a haplogroup in Central Asia during the Neolithic, one would have to compare how many words the Kamkata-viri language (maybe Kamviri dialect) does not share with Sanskrit, Avestan, and European languages.” ref
“If this amount would be much higher than what Sanskrit does not share with Avestan and European languages, then those “not shared” words most likely originated in the R2a associated substrum language, for example, words similar to Sanskrit “मत्स्य (matsya)” (“fish”) and Nuristani Kamviri “oa mâći” (“fish”). In general, there are around 55 non-Indo-European words shared between Sanskrit and Avestan, which might have originated in the language related to the Oxus Civilization. Those 55 words and only 12% of R2a in that region (5% – 10% among other Dravidians in India) disqualify the Dravidian Brahui language as a descendant of a language spoken in the Oxus Civilization.” ref
“The ancient Y-DNA R2a samples predating 1500 BC come mostly from the territory of Turkmenistan and Uzbekistan, so one might suggest that they were related to the Iranic languages (spoken before the arrival of Turkmen and Uzbek languages). This might explain all the differences between Sanskrit and Avestan, but the language of 3500 BCE Turkmenistan and even that of BMAC remains unknown, and in this supposed thesis, the Y‑DNA haplogroup R1a‑Z93 and Sintshta aDNA would be associated with Sanskrit but not with Avestan. Why would then the supposed “R2a associated Iranic languages” be much closer to R1a‑Z93 associated Sanskrit than Sanskrit would be to R1b associated Latin… I have no idea. One would have to prove that Sanskrit is closer to Latin than it is to Avestan or to Nuristani languages.” ref
“To conclude all the speculations… It is not possible to precisely identify a modern language, which would be closest to a language spoken by the people who after 10,000 BCE brought the Y‑DNA haplogroup R2* to Central Asia. This mystery might be solved in the future with a discovery of a language spoken by the Oxus Civilization or the Namazga Culture.” ref
Iran Neolithic
“Demographically complex Near East hints at Anatolian and Indo-Aryan arrival, researchers focused on the three Late Chalcolithic groups with sufficiently large sample size and who are the earliest in time among the LC-LBA groups: ÇamlıbelTarlası_LC (n = 9), İkiztepe_LC (n = 11), and Arslantepe_LC (n = 17). Taking individual estimates from all these individuals together (“Anatolia_LC”), we obtain a robust admixture date estimate of 105 ± 19 generations ago when we use Barcın_N and modern Caucasians as proxies of the source gene pools. Using a generation time of 28 years (Moorjani et al., 2016), this estimate equates to an admixture event ∼3,000 years before the time of the LC-LBA individuals, corresponding to ∼6500 years BCE or around 8,500 years ago.Büyükkaya_EC is the earliest individual in our dataset with a genetic profile similar to the LC-LBA groups within Anatolia.” ref
“Therefore, we also tested a scenario in which the later LC-LBA groups had descended from the same gene pool without further external contribution. f4 (Mbuti, X; Büyükkaya_EC, LC-LBA) suggests that Büyükkaya_EC shares more alleles with the European/Anatolian Farmers (e.g., Boncucklu, LBK, Barcın_N) than LC-LBA groups do. Likewise, most LC-LBA groups cannot be modeled in qpAdm as a sister group with Büyükkaya_EC when Barcın_N is included in the outgroups. The time around 6500–6400 years BCE marks a significant junction in the Anatolian Neolithic because it saw a sudden and massive expansion of sedentary communities into areas that had previously supported no or very few food-producing communities before. Subsequently, in the Southern Caucasus, the abrupt appearance of a Neolithic lifestyle and the introduction of exogenous domesticated animal and plant species ca. 6000 BCE suggests some type of interaction with, and eventually intrusion of Neolithic populations from the neighboring regions, among which Southeastern Anatolia—along with Zagros and the Caspian belt—could be one of the most suitable candidates.” ref
“In the subsequent millennia and until the Late Bronze Age, genetic continuity persisted in North-Central and Eastern Anatolia, which is supported by the genetic similarity of these later populations and the absence of new ancestry sources after the Neolithic period. This contradicts prior hypotheses for population change based on archaeological evidence of intense cultural interactions during this period. For example, the site of İkiztepe on the Turkish Black Sea Coast contains a material culture with strong Balkan affinities, and this has been argued to signify direct contact with populations across the Black Sea, but these contacts do not seem to be accompanied by gene flow.” ref
“Evidence for genetic homogenization across larger geographic distances also comes from the uniparentally inherited Y chromosome lineages. We observe the most common male lineages, J1a, J2a, J2b, and G2a, in all spatiotemporal groups of the region. Alongside the less frequent lineages H2 and T1a, these all form part of the genetic legacy that dates to the Neolithic or was already present in the region during the Upper Paleolithic. In the subsequent millennia and until the Late Bronze Age, genetic continuity persisted in North-Central and Eastern Anatolia, which is supported by the genetic similarity of these later populations and the absence of new ancestry sources after the Neolithic period.” ref
“The paper mentions, in particular, İkiztepe on the Black Sea Coast, which shows a material culture with strong Balkan affinities. This confirms that the available İkiztepe individual from the end-4th millennium BC represented not only the actual community at the time (and was not a recent incomer from the Southern Caucasus), but also that it continued this likely ancestry of Hattic-related North Anatolians homogenized during the previous period, judging by the earliest available sample from the beginning of the millennium. Contrasting with the conclusions about İkiztepe based on a handful of samples, the Arslantepe sampling offers two important cues as to the origin of Anatolian languages and the fate of the lineages brought by their speakers.” ref
“An Arslantepe LC individual, ART038 (ca. 3361-3105 BC), shows hg. R1b-V1636, also highlighted by the paper. The authors, in their outdated comparison to the main haplogroup (R1b-Z2103) reported from Yamnaya and Afanasievo, forget to mention that this same hg. R1b-V1636 was found before in Khvalynsk and in Khvalynsk-related Progress 2, and later in Yamnaya from the Caucasus. ART038 [S150 (H221)] is a young female [sic] from Period VI B1/VI B2 lying on top of stone slabs closing the Royal tomb. Probably sacrificed. Dating of human bone: 3361-3105 cal BCE (4534 ± 27 years ago, MAMS-34112).” ref
“The fact that it has no Steppe ancestry shows how gene flows were easily diluted among a demographically dense Near Eastern population within a few generations. The case of the subsequent Arslantepe EBA period, when Kura-Araxes dominated over the site, is illustrative of a similar process: In PCA and f4-statistics, two individuals dating to this period show excess affinity with populations from the Caucasus and the Pontic steppe compared to their peers, while later Arslantepe_EBA individuals do not share this Caucasian affinity. This implies that the postulated demic interaction must have been transient and of small scale, although the small sample size from Arslantepe_EBA (n = 4) may not be sufficient to detect it. Subtle gene flow is consistent with recent findings from the site, suggesting that the EBA pastoralists occupying the site were more likely well-established local groups moving around the mountains rather than intrusive groups from the Caucasus.” ref
“The only Y-DNA reported from that period is hg. J2a-Z500, which has to be added to the already available Kura-Araxes-related (South Caucasus) G2b-FGC2964, and J1a-Z1842, the three of them thus continuing apparently the previous Near Eastern homogenization period. You can read more about: Ikiztepe and the potential arrival of Proto-Anatolian during the Chalcolithic. A discussion of the complex linguistic landscape of Anatolia, including the North Anatolian family (to which Hattic belongs), and Kura-Araxes as the most likely vector of Hurro-Urartian. Arslantepe and the likely incursion of Early Anatolians, as well as its conquest by non-Indo-European speakers from Kura-Araxes.” ref
Genetic Turnover in the Bronze Age Northern Levant
The Northern Levant, represented by the sites Tell Kurdu, Ebla, and Alalakh, showcases the most noticeable genetic turnover among our four time transects. Within two millennia after the last Middle Chalcolithic Tell Kurdu individual (TellKurdu_MC), the genetic profile of the populations in and around the Amuq valley (Alalakh_MLBA and Ebla_EMBA) changed to largely resemble contemporaneous Anatolians. However, the qpAdm modeling with Büyükkaya_EC suggests that Alalakh_MLBA and Ebla_EMBA are still distinct from the other Anatolian groups with respect to their connection to the ancient Southern Levant. All genetic analyses of Alalakh_MLBA were conducted at the exclusion of one female (ALA019) because of her outlier position in the PCA. Discovered at the bottom of a well, the archaeological and anthropological context suggest that the skeletal remains of this woman, C14-dated to 1568–1511 cal BCE (AMS; 2-sigma), represent an irregular burial with evidence of several healed skeletal traumata (…)” ref
In the Eurasian PCA, ALA019 falls genetically closer to Chalcolithic and Bronze Age individuals from ancient Iran and Turan (present-day area of Iran, Turkmenistan, Uzbekistan, and Afghanistan) (Narasimhan et al., 2019). These populations represent a west-east genetic cline with varying proportions of ancestries related to Barcın_N, Iran_N (Iran Neolithic), and WSHG (hunter-gatherers from Western Siberia). We confirmed the genetic affinity of ALA019 observed in the PCA with an outgroup-f3 test. Other ancient populations from the Caucasus and the Western steppe also produced high affinity but f4(Mbuti, X; Turanx, ALA019) suggest that ALA019 differs from other Turan individuals by occasionally sharing more or less alleles with either Iran_N or WSHG, which agrees with the presence of a genetic cline in this area. In the absence of ancient genomes from nearby regions such as Southern Mesopotamia, the most likely ancestral origin of this individual was somewhere in Eastern Iran or Central Asia.” ref
Interestingly, and even though the sample is female, ALA019 displays an ancestry similar to the region where Bronze Age Near Eastern Indo-Aryanspeakers are supposed to have expanded from. The presence of multiple injuries (potentially dating to her childhood) indicates a hard life, and her ‘burial’ – for lack of a better name – also implies a hard death, but it is difficult to draw any conclusions from a single individual: ALA019 (Square 32.57, Locus 247, AT 15878) is an adult female aged 40-45 years old found at the bottom of a very deep well [hence, dubbed “the Well Lady”]. The remains exhibit the presence of osteoarthritis with eburnation (OA) on the cervical vertebrae between C1 and C2, along with the rare presence of adventitious bursa on lumbar 3 and 4.” ref
“The individual shows evidence of healed trauma on the frontal bone of the skull and two healed fractured ribs (Shafiq, 2020). Enthesophytes were found on both calcaneal bones. (…) She was discovered facedown with her limbs splayed, indicating that she had been carelessly thrown into the well while it was still in use (probably for domestic/craft purposes or for animals). As this individual’s deposition was the result of misadventure, rather than deliberate burial, there are no accompanying grave goods. Dating of human bone: 1625-1511 BCE (3298 ± 23 years ago, MAMS-33687).” ref
“The likely connection of this heavily admixed Turan-like outlier to the arrival of Mitanni Indic suggests, once again, that the expansion of Indo-Iranian languages was a much more complex phenomenon than the Y-DNA bottleneck under R1a-Z93 and Steppe_MLBA visible in Sintashta-Potapovka and continued in the Andronovo horizon. For more on this, see: The initial evolution of Indo-Iranians in Turan and contacts with BMAC. A recent update on how Yamnaya-related peoples replaced Europeans, but admixed heavily as they spread to Asia. The emergence of Indo-Aryan in the Near East and the chronological position of Mitanni Indic.” ref
Early Neolithic genomes from the eastern Fertile Crescent
“Near Eastern genomes from Iran: The genetic composition of populations in Europe changed during the Neolithic transition from hunting and gathering to farming. To better understand the origin of modern populations, Broushaki et al. sequenced ancient DNA from four individuals from the Zagros region of present-day Iran, representing the early Neolithic Fertile Crescent. These individuals unexpectedly were not ancestral to early European farmers, and their genetic structures did not contribute significantly to those of present-day Europeans. These data indicate that a parallel Neolithic transition probably resulted from structured farming populations across southwest Asia.” ref
“Abstract: We sequenced Early Neolithic genomes from the Zagros region of Iran (eastern Fertile Crescent), where some of the earliest evidence for farming is found, and identify a previously uncharacterized population that is neither ancestral to the first European farmers nor has contributed substantially to the ancestry of modern Europeans. These people are estimated to have separated from Early Neolithic farmers in Anatolia some 46,000 to 77,000 years ago and show affinities to modern-day Pakistani and Afghan populations, but particularly to Iranian Zoroastrians. We conclude that multiple, genetically differentiated hunter-gatherer populations adopted farming in southwestern Asia, that components of pre-Neolithic population structure were preserved as farming spread into neighboring regions, and that the Zagros region was the cradle of eastward expansion.” ref
“The earliest evidence for cultivation and stock-keeping is found in the Neolithic core zone of the Fertile Crescent; a region stretching north from the southern Levant through eastern Anatolia and northern Mesopotamia, then east into the Zagros Mountains on the border of modern-day Iran and Iraq. From there, farming spread into surrounding regions, including Anatolia and, later, Europe, southern Asia, and parts of Arabia and North Africa. Whether the transition to agriculture was a homogeneous process across the core zone, or a mosaic of localized domestications, is unknown. Likewise, the extent to which core zone farming populations were genetically homogeneous, or exhibited structure that may have been preserved as agriculture spread into surrounding regions, is undetermined.” ref
“Ancient DNA (aDNA) studies indicate that early Aegean farmers dating to ~6500 to 6000 BCE are the main ancestors of early European farmers, although it is not known if they were predominantly descended from core zone farming populations. We sequenced four Early Neolithic (EN) genomes from Zagros, Iran, including one to 10× mean coverage from a well-preserved male sample from the central Zagros site of Wezmeh Cave [WC1, 7455 to 7082 calibrated years (cal) BCE]. The three other individuals were from Tepe Abdul Hosein and were less well preserved (genome coverage between 0.6 and 1.2×) but are around 10,000 years old, and therefore are among the earliest Neolithic human remains in the world.” ref
“Despite a lack of a clear Neolithic context, the radiocarbon-inferred chronological age and palaeodietary data support WC1 being an early farmer. WC1 bone collagen δ13C and δ15N values are indistinguishable from those of a securely assigned Neolithic individual from Abdul Hosein and consistent with a diet rich in cultivated C3 cereals rather than animal protein. Specifically, collagen from WC1 and Abdul Hosein is 13C depleted compared to those from contemporaneous wild and domestic fauna from this region, which consumed C4 plants. Crucially, WC1 and the Abdul Hosein farmers exhibit very similar genomic signatures.” ref
“The four EN Zagros genomes form a distinct cluster in the first two dimensions of a principal components analysis; they plot closest to modern-day Pakistanis and Afghans and are well separated from European hunter-gatherers (HG) and other Neolithic farmers. In an outgroup f3-test, all four Neolithic Iranian individuals are genetically more similar to each other than to any other prehistoric genome except a Chalcolithic genome from northwestern Anatolia (see below). Despite 14C dates spanning around 1200 years, these data are consistent with all four genomes being sampled from a single eastern Fertile Crescent EN population.” ref
“This observed excess of long segments of reduced heterozygosity could be the result of cultural practices such as consanguinity and endogamy, or demographic constraints such as a recent or ongoing bottleneck. The extent of population genetic structure in Neolithic southwestern Asia has important implications for the origins of farming. High levels of structuring would be expected under a scenario of localized independent domestication processes by distinct populations, whereas low structure would be more consistent with a single population origin of farming or a diffuse homogeneous domestication process, perhaps involving high rates of gene flow across the entire Neolithic core zone. The ancient Zagros individuals show stronger affinities to Caucasus HGs, whereas Neolithic Aegeans showed closer affinities to other European HGs.” ref
“Formal tests of admixture of the form f3 (Neo_Iranian, HG; Anatolia_Neolithic) were all positive with Z-scores above 15.78, indicating that Neolithic northwestern Anatolians did not descend from a population formed by the mixing of Zagros Neolithics and known HG groups. These results suggest that Neolithic populations from northwestern Anatolia and the Zagros descended from distinct ancestral populations. Furthermore, although the Caucasus HGs are genetically closest to EN Zagros individuals, they also share unique ancestry with eastern, western, and Scandinavian European HGs, indicating that they are not the direct ancestors of Zagros Neolithics.” ref
“The significant differences between ancient Iranians, Anatolian/European farmers, and European HGs suggest a pre-Neolithic separation. Assuming a mutation rate of 5 × 10−10 per site per year, the inferred mean split time for Anatolian/European farmers (as represented by Bar8, 4) and European HGs (Loschbour) ranged from 33,000 to 39,000 years ago [combined 95% confidence interval (CI) 15,000 to 61,000 years ago], whereas the preceding divergence of the ancestors of Neolithic Iranians (WC1) occurred 46,000 to 77,000 years ago (combined 95% CI 38,000 to 104,000 years ago). Furthermore, the European HGs were inferred to have an effective population size (Ne) that was ~10 to 20% of either Neolithic farming group, consistent with the ROH and θ ^ analyses.” ref
“Levels of inferred Neanderthal ancestry in WC1 are low, but fall within the general trend described recently in Fu et al. Fu et al. also inferred a basal Eurasian ancestry component in the Caucasus HG sample Satsurblia when examined within the context of a “base model” for various ancient Eurasian genomes dated from ~45,000 to 7,000 years ago. ADMIXTUREGRAPH inferred almost twice as much basal Eurasian ancestry for WC1 as for Satsurblia (62 versus 32%), with the remaining ancestry derived from a population most similar to Ancient North Eurasian (ANE) such as Mal’ta–Buret’ culture. Thus, Neolithic Iranians appear to derive predominantly from the earliest known Eurasian population branching event.” ref
“Chromosome painting” and an analysis of recent haplotype sharing using a Bayesian mixture model revealed that, when compared to 160 to 220 modern groups, WC1 shared a high proportion (>95%) of recent ancestry with individuals from the Middle East, Caucasus, and India. We also compared WC1’s haplotype-sharing profile to that of three high-coverage Neolithic genomes from northwestern Anatolia, Germany (LBK; Stuttgart), and Hungary (NE1; Polgár-Ferenci-hát). Unlike WC1, these Anatolian and European Neolithics shared ~60 to 100% of recent ancestry with modern groups sampled from southern Europe.” ref
“Recent haplotype sharing between each modern group and ancient Neolithic genomes from Iran (WC1) and Europe (LBK, NE1), HG genomes sampled from Luxembourg (Loschbour) and the Caucasus (KK1; Kotias), a 4500-year-old genome from Ethiopia (Mota) and Ust’-Ishim, and a 45,000-year-old genome from Siberia. Modern groups from south, central, and northwestern Europe shared haplotypes predominantly with European Neolithic samples LBK and NE1, and European HGs, whereas modern Near and Middle Eastern, as well as southern Asian samples, had higher sharing with WC1. Modern Pakistani, Iranian, Armenian, Tajikistani, Uzbekistani, and Yemeni samples were inferred to share >10% of haplotypes with WC1. This was true even when modern groups from neighboring geographic regions were added as potential ancestry surrogates. Iranian Zoroastrians had the highest inferred sharing with WC1 out of all modern groups. Consistent with this, outgroup f3 statistics indicate that Iranian Zoroastrians are the most genetically similar to all four Neolithic Iranians, followed by other modern Iranians (Fars), Balochi (southeastern Iran, Pakistan, and Afghanistan), Brahui (Pakistan and Afghanistan), Kalash (Pakistan), and Georgians. Interestingly, WC1 most likely had brown eyes, relatively dark skin, and black hair, although Neolithic Iranians carried reduced pigmentation-associated alleles in several genes and derived alleles at 7 of the 12 loci showing the strongest signatures of selection in ancient Eurasians. Although there is a strong Neolithic component in these modern south Asian populations, simulation of allele sharing rejected full population continuity under plausible ancestral population sizes, indicating some population turnover in Iran since the Neolithic.” ref
“While Early Neolithic samples from eastern and western southwest Asia differ conspicuously, comparisons to genomes from Chalcolithic Anatolia and Iron Age Iran indicate a degree of subsequent homogenization. Kumtepe6, a ~6750-year-old genome from northwestern Anatolia, was more similar to Neolithic Iranians than to any other non-Iranian ancient genome. Furthermore, our male Iron Age genome (971 to 832 BCE) from Tepe Hasanlu in northwestern Iran shares greatest similarity with Kumtepe6 even when compared to Neolithic Iranians (table S20). We inferred additional non-Iranian or non-Anatolian ancestry in F38 from sources such as European Neolithics and even post-Neolithic Steppe populations (table S20). Consistent with this, F38 carried a N1a subclade mitochondrial DNA (mtDNA), which is common in early European and northwestern Anatolian farmers. In contrast, his Y chromosome belongs to subhaplogroup R1b1a2a2, also found in five Yamnaya individuals and in two individuals from the Poltavka culture. These patterns indicate that post-Neolithic homogenization in southwestern Asia involved substantial bidirectional gene flow between the east and west of the region, as well as possible gene flow from the Steppe.” ref
“Migration of people associated with the Yamnaya culture has been implicated in the spread of Indo-European languages, and some level of Near Eastern ancestry was previously inferred in southern Russian pre-Yamnaya populations. However, our analyses suggest that Neolithic Iranians were unlikely to be the main source of Near Eastern ancestry in the Steppe population (table S20) and that this ancestry in pre-Yamnaya populations originated primarily in the west of southwest Asia. Researchers also inferred shared ancestry between Steppe and Hasanlu Iron Age genomes that was distinct from EN Iranians. In addition, modern Middle Easterners and South Asians appear to possess mixed ancestry from ancient Iranian and Steppe populations (tables S19 and S20). However, Steppe-related ancestry may also have been acquired indirectly from other sources, and it is not clear if this is sufficient to explain the spread of Indo-European languages from a hypothesized Steppe homeland to the region where Indo-Iranian languages are spoken today. Yet, the affinities of Zagros Neolithic individuals to modern populations of Pakistan, Afghanistan, Iran, and India is consistent with a spread of Indo-Iranian languages, or of Dravidian languages (which includes Brahui), from the Zagros into southern Asia, in association with farming.” ref
“The Neolithic transition in southwest Asia involved the appearance of different domestic species, particularly crops, in different parts of the Neolithic core zone, with no single center. Early evidence of plant cultivation and goat management between the 10th and the 8th millennium BCE highlights the Zagros as a key region in the Neolithization process. Given the evidence of domestic species movement from east to west across southwest Asia, it is surprising that EN human genomes from the Zagros are not closely related to those from northwestern Anatolia and Europe. Instead, they represent a previously undescribed Neolithic population. Our data show that the chain of Neolithic migration into Europe does not reach back to the eastern Fertile Crescent, also raising questions about whether intermediate populations in southeastern and Central Anatolia form part of this expansion. Nevertheless, it seems probable that the Zagros region was the source of an eastern expansion of the southwestern Asian domestic plant and animal economy. Our inferred persistence of ancient Zagros genetic components in modern-day South Asians lends weight to a strong demic component to this expansion.” ref
The genetics of an early Neolithic pastoralist from the Zagros, Iran
“Abstract: The agricultural transition profoundly changed human societies. We sequenced and analyzed the first genome (1.39x) of an early Neolithic woman from Ganj Dareh, in the Zagros Mountains of Iran, a site with early evidence for an economy based on goat herding, ca. 10,000 years ago. We show that Western Iran was inhabited by a population genetically most similar to hunter-gatherers from the Caucasus, but distinct from the Neolithic Anatolian people who later brought food production into Europe. The inhabitants of Ganj Dareh made little direct genetic contribution to modern European populations, suggesting those of the Central Zagros were somewhat isolated from other populations of the Fertile Crescent. Runs of homozygosity are of a similar length to those from Neolithic farmers and shorter than those of Caucasus and Western Hunter-Gatherers, suggesting that the inhabitants of Ganj Dareh did not undergo the large population bottleneck suffered by their northern neighbors. While some degree of cultural diffusion between Anatolia, Western Iran, and other neighboring regions is possible, the genetic dissimilarity between early Anatolian farmers and the inhabitants of Ganj Dareh supports a model in which Neolithic societies in these areas were distinct.” ref
Ancient Migratory Events in the Middle East: New Clues from the Y-Chromosome Variation of Modern Iranians
“Abstract: Knowledge of high-resolution Y-chromosome haplogroup diversification within Iran provides an important geographic context regarding the spread and compartmentalization of male lineages in the Middle East and southwestern Asia. At present, the Iranian population is characterized by an extraordinary mix of different ethnic groups speaking a variety of Indo-Iranian, Semitic, and Turkic languages. Despite these features, only a few studies have investigated the multiethnic components of the Iranian gene pool. In this survey, 938 Iranian male DNAs belonging to 15 ethnic groups from 14 Iranian provinces were analyzed for 84 Y-chromosome biallelic markers and 10 STRs. The results show an autochthonous but non-homogeneous ancient background mainly composed by J2a sub-clades with different external contributions. The phylogeography of the main haplogroups allowed the identification of post-glacial and Neolithic expansions toward western Eurasia but also recent movements towards the Iranian region from western Eurasia (R1b-L23), Central Asia (Q-M25), Asia Minor (J2a-M92) and southern Mesopotamia (J1-Page08). In spite of the presence of important geographic barriers (Zagros and Alborz mountain ranges and the Dasht-e Kavir and Dash-e Lut deserts), which may have limited gene flow, AMOVA analysis revealed that language, in addition to geography, has played an important role in shaping the nowadays Iranian gene pool. Overall, this study provides a portrait of the Y-chromosomal variation in Iran, useful for depicting a more comprehensive history of the peoples of this area as well as for reconstructing ancient migration routes. In addition, our results evidence the important role of the Iranian plateau as a source and recipient of gene flow between culturally and genetically distinct populations.” ref
“The Middle Eastern region had a central role in human evolution. It has been a passageway for Homo sapiens between Africa and the rest of Asia and, in particular, the first region of the Asian continent occupied by modern humans. This area was also one of the regions where agriculture began during the Neolithic period, in particular in the Fertile Crescent, from which it spread westwards and eastwards. Different pre-historic sites across the Iranian plateau point to the existence of ancient cultures and urban settlements in the sixth millennium BP, perhaps even some centuries earlier than the earliest civilizations in nearby Mesopotamia. The proto-Iranian language first emerged following the separation of the Indo-Iranian branch from the Indo-European language family. Proto-Iranian tribes from Central Asian steppes arrived on the Iranian plateau in the fifth and fourth millennium years ago, settled as nomads, and further separated into different groups. By the third millennium BP, Cimmerians, Sarmatians, and Alans populated the steppes North of the Black Sea, while Medes, Persians, Bactrians, and Parthians occupied the western part of the Iranian plateau. Other tribes began to settle on the eastern edge, as far East as on the mountainous frontier of north-western Indian subcontinent and into the area which is now Baluchistan.” ref
“Nowadays, Iranian territory has been occupied by Medes (Maad) in the central and north-western regions, Persians (Paars) in the south-western region, and Parthians (Parthav) in the north-eastern and eastern regions of the country. In the 6th century BCE Cyrus the Great founded the Achaemenid Empire (the first Persian Empire), which started in South Iran and spread from Libya to Anatolia and Macedonia, encompassing an extraordinary ethno-cultural diversity. This widespread empire collapsed after two centuries (towards the end of the 4th century BCE) on account of Alexander the Great. In the 2nd century BC, north-eastern Persia was invaded by the Parthians who founded an empire extending from the Euphrates to Afghanistan. Because of its location on the Silk Road, connecting the Roman Empire and the Han Dynasty in China, it quickly became a center of trade and commerce. The Parthians were succeeded by the Sassanid Empire, one of the most important and influential historical periods of Persia. Afterwards, Iran was invaded by several populations, such as the Arabs, Mongols, and Ottoman Turks. The Muslim conquest of Persia in 637 CE led to the introduction of Islam, with the consequent decline of the Zoroastrian religion, which still survives in some communities in different parts of Iran, especially in Tehran and Yazd.” ref
“This continuous invasion of populations with different origins and cultures created an interesting mix of different ethnic groups speaking a variety of Indo-Iranian, Semitic, and Turkic languages and encompassing Arabs, Armenians, Assyrians, Azeris, Baluchs, Bandaris, Gilaks, Kurds, Lurs, Mazandarani, Persians, Qeshm people, Turkmens, Zoroastrians and a group of so-called Afro-Iranians, which might be the result of the slave trade with Zanzibar. Despite the great potentiality of this genetic scenario in providing useful information to reconstruct traces of ancient migrations, only a few studies have investigated the multi-ethnic components of the Iranian gene pool. In order to shed some light on the genetic structure of the Iranian population as well as on the expansion patterns and population movements that affected this region, the Y-chromosomes of 938 Iranians, representative of the majority of the provinces and ethnic groups in Iran, were examined at an unprecedented level of resolution.” ref
“The analysis of 88 Y-chromosome bi-allelic markers in 938 subjects belonging to 15 ethnic groups from 14 Iranian provinces allowed the identification of 65 different Y-chromosome lineages (Table 1 and Figure S1). They belong to 15 main haplogroups (B, C, D, E, F, G, H, I, J, L, N, O, Q, R and T) the most frequent of which are J (31.4%), R (29.1%), G (11.8%) and E (9.2%), with great differences (disregarding those relative to samples smaller than 20 subjects) in frequencies and sub-haplogroups observed among provinces and ethnic groups (Figure 1). On the whole, the Iranian population is characterized by very high haplogroup diversity (0.952): the maximum value being observed in the Persians of Fars (0.962) and the minimum in the Arabs of Khuzestan (0.883) and the Turkmen of Golestan (0.821).” ref
“Haplogroup J is predominant in Iran where both its sub-clades, J2-M172 and J1-M267, are observed. Its highest frequencies are registered in the populations located along the south-western shores of the Caspian Sea and along the Zagros Mountains ridge. Exceptionally high is the frequency observed in the Baluchi of Sistan Baluchestan, in agreement with their likely Caspian Sea origin.” ref
“J1-M267 does not exceed 10% in the majority of the Iranian samples examined, with higher values only in Fars (11.4%), Zoroastrians from Yazd (11.7%), Gilan (12.5%), Assyrians from Azerbaijan (17.9%) and Khuzestan (33.4%). The proportion of the two sub-lineages, J1-Page08 and J1-M267*, is highly variant, being J1-M267* almost restricted to north-western Iranian groups and J1-Page08 mainly observed in populations living below the Dasht-e Kevir and Dasht-e Lut desert area, (approximately latitude 30°N). It reaches a frequency of 31.6% in the Arab group from Khuzestan at the border with southern Iraq. J2-M172 is the main Iranian haplogroup (22.5%), almost entirely (92.9%) represented by J2a-M410 sub-clades.” ref
“The majority of the M410 chromosomes are J2a-Page55 and mainly represented by its main sub-clades M530, M47, and M67. In particular, the recently described J2a-M530 shows high frequencies in the Zoroastrians of Yazd (17.6%) and Tehran (15.4%), and in the Persians of Yazd (17.0%). J2a-M47 reaches frequencies higher than 5% in the Zoroastrians of Yazd (8.8%), in Mazandaran, Khuzestan, and Fars (∼7%), while it is absent in the Assyrians of Azerbaijan Gharbi and Tehran, in Sistan Baluchestan and in Hormozgan (except for the Qeshm group). J2a-M92 was observed in Sistan Baluchestan (12.5%) while the paragroup J2a-M67* was observed mainly in the Armenians of Tehran (8.8%). J2a-M68, previously reported in the neighboring Iraqi population, was not observed in Iran. As for the paragroups, J2a-M410* represents 2.8% of the total sample with ∼7% of frequency in Khuzestan, Mazandaran, and Khorasan, whereas J2a-Page55*, observed at 6.6% in central Anatolia, accounts for 4.8% of the Iranian sample. J2-M172*, recently described in the neighboring Iraqi Marsh Arabs (3.5%), characterizes one subject from Khuzestan (1.8%).” ref
“Haplogroup R in Iran is mainly represented by the R1 sub-lineages R1a-M198 and R1b-M269, whereas R2-M124 was observed only in 2.8% of the total sample. All the R1a Y chromosomes belong to the M198* paragroup with frequencies ranging from 0% to 25%. Indeed neither the “European” M458 nor the “Pakistani” M434 have been observed in our samples. Haplogroup R1b-M269 shows its highest frequency in the Assyrians (29.2%, averaged on Tehran and Azerbaijan Gharbi groups). High values are also observed in the Armenians from Tehran and in Lorestan (both with ∼24%). With the exception of five chromosomes belonging to the paragroup R1b-M269* and three chromosomes clustering in the “European” sub-haplogroup R1b-M412, all the M269 Y chromosomes belong to the R1b-L23 clade.” ref
“Haplogroup G is observed in this survey as G1-M285 and G2a-P15. G1-M285, previously described in the Iranian population, accounts only for 1.8% of the present Iranian sample. G2a-P15 is the most frequent sub-clade characterizing 9.1% of the total sample, with incidences ranging from 0% in Sistan Baluchestan to 19.3% in the Arabs of Khuzestan. Interestingly, the majority (74.7%) of the G2a-P15 Y chromosomes belong to the paragroups G2a-P15* and G2a-P303*.” ref
“Haplogroup E in Iran is mainly represented by the E1-M123 (3.7%) and E1b-M78 (3.0%) branches. The first is almost entirely characterized by its sub-lineage M34 and reaches its highest incidence (13.6%) in Kurdistan. The second is present as E1b-M78* in Lorestan (9.8%), E1b-V13 (5.9%) and E1b-V22 (2.9%) in the Zoroastrians of Yazd. It is worth noting the presence of individuals carrying African-specific haplogroups (three belonging to E2-M75 and 17 to E1b-M2) in South-East Iran (Hormozgan and Sistan Baluchestan), whereas the North-East African E1b-M81 is not observed.” ref
Evidence of Late Glacial expansions from a Near Eastern Y-chromosome reservoir
“It is known that in parts of the Near East, such as the Levant and Asia Minor, populations persisted throughout the last glaciation but no archaeological evidence for a Near Eastern Late Glacial expansions has till now been discovered. Recently, thanks to the recalibration of the mitochondrial DNA (mtDNA) clock, signals of Near Eastern dispersals towards Europe in the Late Glacial (from 12,000–19,000 years ago) emerged from complete mitochondrial genome analysis of haplogroups J and T, previously associated only with the Neolithic diffusion. Although the Y-chromosome molecular clock is far from reaching the mtDNA level of accuracy, evidences of Late Glacial dispersals from the Middle East are provided by the large number of deep rooting lineages (rare elsewhere), from which diverged different branches that underwent Neolithic expansions. Accordingly, Y chromosomes F-M89* and IJ-M429* were observed in the Iranian plateau: the first represents the ancestral state of the main Euro-Asiatic haplogroups while the second probably moved toward southeast Europe sometime before the Last Glacial Maximum where it differentiated into the “western Eurasian” haplogroup I. Similarly, basal lineages of the “Middle Eastern” haplogroup J (J1-M267* and different J2a lineages: J2-M172*, J2a-M410* and J2a-Page55*) and of haplogroups G (G2-P287*, G2-P15* and G2-P303*) and R (R1b-M269*) were also observed. Their frequency and variance distributions suggest a Mesolithic Middle Eastern origin/presence (Figure 1, Tables S2, S3, S4, S5 and S7) of these Y chromosomes supporting the role of the Middle East as a genetic reservoir for Late Glacial expansions and subsequent Neolithic dispersals southwards and westwards into South-East Europe.” ref
“J1-M267* shows high variance in the Middle Eastern region including Eastern Turkey, North-West Iraq, and North-West Iran (Gilan – Mazandaran, Table S2), where probably originated 26,300 ±8.2 years ago (Table S7) and then migrated westwards up to the Balkans and the Italian Peninsula and southwards as far as in Saudi Arabia and Ethiopia. The network of the M267* haplotypes (Figures 2 and S2) confirms the previously described non-star-like substructure, enlightening a recent expansion (5,500 ±2.9 years ago, Table S7) of the cluster characterized by the DYS388-13 and DYS390-23 repeats including North-East Turkish and Assyrian (from Turkey, Iraq and Iran) Y-chromosomes. This cluster also harbours virtually all the M267* Marsh Arab Y chromosomes supporting the previously proposed origin in northern Mesopotamia for the Iraqi Marsh Arabs. However, only a further subdivision of this paragroup will allow a better understanding of times and ways of migrations marked by the M267* Y chromosomes.
“Among the different J2a haplogroups, J2a-M530 is the most informative as for ancient dispersal events from the Iranian region. This lineage probably originated in Iran where it displays its highest frequency and variance in Yazd and Mazandaran (Figure 2). Taking into account its microsatellite variation and age estimates along its distribution area (Tables S3 and S7), it is likely that its diffusion could have been triggered by the Euroasiatic climatic amelioration after the Last Glacial Maximum and later increased by agriculture spread from Turkey and Caucasus towards southern Europe. The high variance observed in the Italian Peninsula is probably the result of stratifications of subsequent migrations and/or of the presence of sub-lineages not yet identified. Of interest in the M530 network (Figures 2 and S3) is the presence of a lateral branch that is characterized by a DYS391 repeat number equal to 9. Differently from previous observations, this branch is not restricted to Anatolian Greek samples being shared with different eastern Mediterranean coastal populations. The M530 diffusion pattern seems to be also shared by the paragroups J2a-M410* and J2a-PAGE55*. In addition, the variance distribution of the rare R1b-M269* Y chromosomes, displaying decreasing values from Iran, Anatolia and the western Black Sea coastal region, is also suggestive of a westward diffusion from the Iranian plateau, although more complex scenarios can be still envisioned because of its non-star like structure.” ref
“Another lineage potentially informative in revealing pre-Neolithic dispersals from the Middle East towards Europe is J2a-M67*. It is characterized by a wide distribution, including European, North-African and Near Eastern Y chromosomes, without virtually going beyond Afghanistan and Pakistan. Its variance distribution identifies different frequency peaks in Iran, the Levant, Cyprus, Crete and Central Italy (Figure 2). The network (Figures 2 and S4), which appears to be complex reflecting internal heterogeneity, includes three most frequent, one step related, haplotypes harboring chromosomes from different populations, few common haplotypes (within population sub-sets) and a wide number of singleton haplotypes. Expansion events are clearly identified in the Levant and the Anatolia/Caucasus/southern Balkan regions from where the M67* spread towards southern Europe. Differently, no sign of J2a-M67* expansion is registered in other areas at high variance such as Iran (15,800 ±4.0 years ago), Cyprus (14,800 ±4.0 years ago), Central Italy (13,200 ±4.2 years ago) and Crete (12,900 ±4.5 years ago) (Table S7) where the majority of the observed haplotypes are rare and occupy a peripheral position in the network. Thus, while the high M67* variance in Central Italy is likely due to a stratification of seaborne migrations of Middle Eastern/Asia Minor peoples, the diversification observed in Iran and the Aegean Islands can be explained by a first Near Eastern, and possibly Anatolian, diffusion of the lineage followed by a Levantine expansion.” ref
Haplogroup R1a and the diffusion of Indo-European languages
“The diffusion of the Iranian branch of Indo-European languages whose origin is generally attributed to a western Asian region which includes Anatolia, the South Caucasus and the North Pontic-Caspian area; has been linked by numerous authors to the R1a haplogroup dispersal. However, in spite of the recent dissection of this haplogroup, none of the identified sub-branches support a patrilineal gene flow from western Eurasia through southern Asia ascribable to the diffusion of Indo-European languages. Accordingly, the present analysis of the Iranian R1a Y-chromosomes does not provide useful information to disentangle this issue. Indeed, the Iranian Y-chromosomes, as the majority of the European and virtually all the Asian ones, are still part of the unresolved paragroup R1a-M198* and harbour haplotypes shared by both European and Asian Y chromosomes.” ref
Recent gene flows from neighbouring populations
Traces of recent gene flows from Arab countries and Anatolia are revealed in the Iranian Y-chromosome gene pool by the presence of the well-resolved sub-haplogroups J1-Page08 and J2-M92, respectively. The “Arab” J1-Page08, likely originated in the region at the border between south-eastern Turkey and North Iraq, underwent an important Neolithic expansion in the southern countries of the Middle East and represents the most important haplogroup in the modern populations of the Arabian Peninsula and North Africa. This lineage is observed at an averaged frequency of 6% in Iran, reaching a value in the Khuzestan Arabs (31.6%, Table 1), which is comparable to that observed in the neighboring Iraqi population. J2a-M92 is a well-defined J2a-M67 sub-lineage, with a distribution restricted to Asia Minor, the Balkans, and the north-eastern Mediterranean coasts. Frequency and variance maps make plausible an origin in north-western Turkey, where the highest variance is registered, and a subsequent migration to the Balkans and then to the Italian Peninsula. In Iran it is sporadically observed with the only exception of Sistan Baluchestan where it reaches an incidence of 12.5%. According to the age estimate (1,300 ±1.3 years ago, Table S7) of the microsatellite variation associated to J2a-M92, its presence in Iran is ascribable to recent gene flow.” ref



ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref
Late Pleistocene human genome suggests a local origin for the first farmers of central Anatolia
“Anatolia was home to some of the earliest farming communities. It has been long debated whether a migration of farming groups introduced agriculture to central Anatolia. Here, we report the first genome-wide data from a 15,000-year-old Anatolian hunter-gatherer and from seven Anatolian and Levantine early farmers. We find high genetic continuity (~80–90%) between the hunter-gatherers and early farmers of Anatolia and detect two distinct incoming ancestries: an early Iranian/Caucasus related one and a later one linked to the ancient Levant. Finally, we observe a genetic link between southern Europe and the Near East predating 15,000 years ago. Our results suggest a limited role of human migration in the emergence of agriculture in central Anatolia.” ref
Agriculture in Iran
“Agriculture has a long history and tradition in Iran. As early as 10,000 BCE, the earliest known domestication of the goat had taken place in the Iranian plateau. By 5,000 BCE, wine was being fermented in Iran, and by as early as 7th century CE, the windmill had been invented in Persia for the first time in history.” ref





Proto-Semitic dates to ca. 4500–3500 BCE or around 6,500 to 5,500 years ago.


The lunar-crescent-shaped monument is massive at about 14,000 cubic meters (500,000 cubic feet) and has a length of about 150 meters (492 feet), 3050 to 2650 BCE, or around 5,050 to 4,650 years ago.
The 4,500-year-old chalk earthworks monument at Durrington Walls, the super-henge is the largest henge in Britain, has a row of megaliths, with 90 standing stones, some 15 feet tall, is less than 2 miles from Stonehenge.

ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref
By day the LORD went ahead of them in a pillar of cloud to guide them on their way and by night in a pillar of fire to give them light, so that they could travel by day or night.
- By day the “Bible God” was in a cloud pillar.
- By night the “Bible God” was in a fire pillar.
- Did a 4,520–4,420-year-old Volcano In Turkey Inspire the Bible God?


.png){kind=link}
.PNG){kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
.png){kind=link}

My favorite “Graham Hancock” Quote?
“In what archaeologists have studied, yes, we can say there is NO Evidence of an advanced civilization.” – (Time 1:27) Joe Rogan Experience #2136 – Graham Hancock & Flint Dibble
Help the Valentine fight against pseudoarchaeology!!!
In a world of “Hancocks” supporting evidence lacking claims, be a “John Hoopes” supporting what evidence explains.
#SupportEvidenceNotWishfullThinking
Graham Hancock: @Graham__Hancock
John Hoopes: @KUHoopes

People don’t commonly teach religious history, even that of their own claimed religion. No, rather they teach a limited “pro their religion” history of their religion from a religious perspective favorable to the religion of choice.

Do you truly think “Religious Belief” is only a matter of some personal choice?
Do you not see how coercive one’s world of choice is limited to the obvious hereditary belief, in most religious choices available to the child of religious parents or caregivers? Religion is more commonly like a family, culture, society, etc. available belief that limits the belief choices of the child and that is when “Religious Belief” is not only a matter of some personal choice and when it becomes hereditary faith, not because of the quality of its alleged facts or proposed truths but because everyone else important to the child believes similarly so they do as well simply mimicking authority beliefs handed to them. Because children are raised in religion rather than being presented all possible choices but rather one limited dogmatic brand of “Religious Belief” where children only have a choice of following the belief as instructed, and then personally claim the faith hereditary belief seen in the confirming to the belief they have held themselves all their lives. This is obvious in statements asked and answered by children claiming a faith they barely understand but they do understand that their family believes “this or that” faith, so they feel obligated to believe it too. While I do agree that “Religious Belief” should only be a matter of some personal choice, it rarely is… End Hereditary Religion!

Animism: Respecting the Living World by Graham Harvey
“How have human cultures engaged with and thought about animals, plants, rocks, clouds, and other elements in their natural surroundings? Do animals and other natural objects have a spirit or soul? What is their relationship to humans? In this new study, Graham Harvey explores current and past animistic beliefs and practices of Native Americans, Maori, Aboriginal Australians, and eco-pagans. He considers the varieties of animism found in these cultures as well as their shared desire to live respectfully within larger natural communities. Drawing on his extensive casework, Harvey also considers the linguistic, performative, ecological, and activist implications of these different animisms.” ref

We are like believing machines we vacuum up ideas, like Velcro sticks to almost everything. We accumulate beliefs that we allow to negatively influence our lives, often without realizing it. Our willingness must be to alter skewed beliefs that impend our balance or reason, which allows us to achieve new positive thinking and accurate outcomes.

My thoughts on Religion Evolution with external links for more info:
- (Pre-Animism Africa mainly, but also Europe, and Asia at least 300,000 years ago), (Pre-Animism – Oxford Dictionaries)
- (Animism Africa around 100,000 years ago), (Animism – Britannica.com)
- (Totemism Europe around 50,000 years ago), (Totemism – Anthropology)
- (Shamanism Siberia around 30,000 years ago), (Shamanism – Britannica.com)
- (Paganism Turkey around 12,000 years ago), (Paganism – BBC Religion)
- (Progressed Organized Religion “Institutional Religion” Egypt around 5,000 years ago), (Ancient Egyptian Religion – Britannica.com)
- (CURRENT “World” RELIGIONS after 4,000 years ago) (Origin of Major Religions – Sacred Texts)
- (Early Atheistic Doubting at least by 2,600 years ago) (History of Atheism – Wikipedia)
“Religion is an Evolved Product” and Yes, Religion is Like Fear Given Wings…
Atheists talk about gods and religions for the same reason doctors talk about cancer, they are looking for a cure, or a firefighter talks about fires because they burn people and they care to stop them. We atheists too often feel a need to help the victims of mental slavery, held in the bondage that is the false beliefs of gods and the conspiracy theories of reality found in religions.
Understanding Religion Evolution:
- Pre-Animism (at least 300,000 years ago)
- Animism (Africa: 100,000 years ago)
- Totemism (Europe: 50,000 years ago)
- Shamanism (Siberia: 30,000 years ago)
- Paganism (Turkey: 12,000 years ago)
- Progressed organized religion (Egypt: 5,000 years ago), (Egypt, the First Dynasty 5,150 years ago)
- CURRENT “World” RELIGIONS (after 4,000 years ago)
- Early Atheistic Doubting (at least by 2,600 years ago)
“An Archaeological/Anthropological Understanding of Religion Evolution”
It seems ancient peoples had to survived amazing threats in a “dangerous universe (by superstition perceived as good and evil),” and human “immorality or imperfection of the soul” which was thought to affect the still living, leading to ancestor worship. This ancestor worship presumably led to the belief in supernatural beings, and then some of these were turned into the belief in gods. This feeble myth called gods were just a human conceived “made from nothing into something over and over, changing, again and again, taking on more as they evolve, all the while they are thought to be special,” but it is just supernatural animistic spirit-belief perceived as sacred.
Quick Evolution of Religion?
Pre-Animism (at least 300,000 years ago) pre-religion is a beginning that evolves into later Animism. So, Religion as we think of it, to me, all starts in a general way with Animism (Africa: 100,000 years ago) (theoretical belief in supernatural powers/spirits), then this is physically expressed in or with Totemism (Europe: 50,000 years ago) (theoretical belief in mythical relationship with powers/spirits through a totem item), which then enlists a full-time specific person to do this worship and believed interacting Shamanism (Siberia/Russia: 30,000 years ago) (theoretical belief in access and influence with spirits through ritual), and then there is the further employment of myths and gods added to all the above giving you Paganism (Turkey: 12,000 years ago) (often a lot more nature-based than most current top world religions, thus hinting to their close link to more ancient religious thinking it stems from). My hypothesis is expressed with an explanation of the building of a theatrical house (modern religions development). Progressed organized religion (Egypt: 5,000 years ago) with CURRENT “World” RELIGIONS (after 4,000 years ago).
Historically, in large city-state societies (such as Egypt or Iraq) starting around 5,000 years ago culminated to make religion something kind of new, a sociocultural-governmental-religious monarchy, where all or at least many of the people of such large city-state societies seem familiar with and committed to the existence of “religion” as the integrated life identity package of control dynamics with a fixed closed magical doctrine, but this juggernaut integrated religion identity package of Dogmatic-Propaganda certainly did not exist or if developed to an extent it was highly limited in most smaller prehistoric societies as they seem to lack most of the strong control dynamics with a fixed closed magical doctrine (magical beliefs could be at times be added or removed). Many people just want to see developed religious dynamics everywhere even if it is not. Instead, all that is found is largely fragments until the domestication of religion.
Religions, as we think of them today, are a new fad, even if they go back to around 6,000 years in the timeline of human existence, this amounts to almost nothing when seen in the long slow evolution of religion at least around 70,000 years ago with one of the oldest ritual worship. Stone Snake of South Africa: “first human worship” 70,000 years ago. This message of how religion and gods among them are clearly a man-made thing that was developed slowly as it was invented and then implemented peace by peace discrediting them all. Which seems to be a simple point some are just not grasping how devastating to any claims of truth when we can see the lie clearly in the archeological sites.
I wish people fought as hard for the actual values as they fight for the group/clan names political or otherwise they think support values. Every amount spent on war is theft to children in need of food or the homeless kept from shelter.
Here are several of my blog posts on history:
- To Find Truth You Must First Look
- (Magdalenian/Iberomaurusian) Connections to the First Paganists of the early Neolithic Near East Dating from around 17,000 to 12,000 Years Ago
- Natufians: an Ancient People at the Origins of Agriculture and Sedentary Life
- Possible Clan Leader/Special “MALE” Ancestor Totem Poles At Least 13,500 years ago?
- Jewish People with DNA at least 13,200 years old, Judaism, and the Origins of Some of its Ideas
- Baltic Reindeer Hunters: Swiderian, Lyngby, Ahrensburgian, and Krasnosillya cultures 12,020 to 11,020 years ago are evidence of powerful migratory waves during the last 13,000 years and a genetic link to Saami and the Finno-Ugric peoples.
- The Rise of Inequality: patriarchy and state hierarchy inequality
- Fertile Crescent 12,500 – 9,500 Years Ago: fertility and death cult belief system?
- 12,400 – 11,700 Years Ago – Kortik Tepe (Turkey) Pre/early-Agriculture Cultic Ritualism
- Ritualistic Bird Symbolism at Gobekli Tepe and its “Ancestor Cult”
- Male-Homosexual (female-like) / Trans-woman (female) Seated Figurine from Gobekli Tepe
- Could a 12,000-year-old Bull Geoglyph at Göbekli Tepe relate to older Bull and Female Art 25,000 years ago and Later Goddess and the Bull cults like Catal Huyuk?
- Sedentism and the Creation of goddesses around 12,000 years ago as well as male gods after 7,000 years ago.
- Alcohol, where Agriculture and Religion Become one? Such as Gobekli Tepe’s Ritualistic use of Grain as Food and Ritual Drink
- Neolithic Ritual Sites with T-Pillars and other Cultic Pillars
- Paganism: Goddesses around 12,000 years ago then Male Gods after 7,000 years ago
- First Patriarchy: Split of Women’s Status around 12,000 years ago & First Hierarchy: fall of Women’s Status around 5,000 years ago.
- Natufians: an Ancient People at the Origins of Agriculture and Sedentary Life
- J DNA and the Spread of Agricultural Religion (paganism)
- Paganism: an approximately 12,000-year-old belief system
- Paganism 12,000 years old: related to “Anarchism and Socialism” (Pre-Capitalism)
- Shaman burial in Israel 12,000 years ago and the Shamanism Phenomena
- Need to Mythicized: gods and goddesses
- 12,000 – 7,000 Years Ago – Paleo-Indian Culture (The Americas)
- 12,000 – 2,000 Years Ago – Indigenous-Scandinavians (Nordic)
- Norse did not wear helmets with horns?
- Pre-Pottery Neolithic Skull Cult around 11,500 to 8,400 Years Ago?
- 10,400 – 10,100 Years Ago, in Turkey the Nevail Cori Religious Settlement
- 9,000-6,500 Years Old Submerged Pre-Pottery/Pottery Neolithic Ritual Settlements off Israel’s Coast
- Catal Huyuk “first religious designed city” around 9,500 to 7,700 years ago (Turkey)
- Cultic Hunting at Catal Huyuk “first religious designed city”
- Special Items and Art as well as Special Elite Burials at Catal Huyuk
- New Rituals and Violence with the appearance of Pottery and People?
- Haplogroup N and its related Uralic Languages and Cultures
- Ainu people, Sámi people, Native Americans, the Ancient North Eurasians, and Paganistic-Shamanism with Totemism
- Ideas, Technology and People from Turkey, Europe, to China and Back again 9,000 to 5,000 years ago?
- First Pottery of Europe and the Related Cultures
- 9,000 years old Neolithic Artifacts Judean Desert and Hills Israel
- 9,000-7,000 years-old Sex and Death Rituals: Cult Sites in Israel, Jordan, and the Sinai
- 9,000-8500 year old Horned Female shaman Bad Dürrenberg Germany
- Neolithic Jewelry and the Spread of Farming in Europe Emerging out of West Turkey
- 8,600-year-old Tortoise Shells in Neolithic graves in central China have Early Writing and Shamanism
- Swing of the Mace: the rise of Elite, Forced Authority, and Inequality begin to Emerge 8,500 years ago?
- Migrations and Changing Europeans Beginning around 8,000 Years Ago
- My “Steppe-Anatolian-Kurgan hypothesis” 8,000/7,000 years ago
- Around 8,000-year-old Shared Idea of the Mistress of Animals, “Ritual” Motif
- Pre-Columbian Red-Paint (red ochre) Maritime Archaic Culture 8,000-3,000 years ago
- 7,522-6,522 years ago Linear Pottery culture which I think relates to Arcane Capitalism’s origins
- Arcane Capitalism: Primitive socialism, Primitive capital, Private ownership, Means of production, Market capitalism, Class discrimination, and Petite bourgeoisie (smaller capitalists)
- 7,500-4,750 years old Ritualistic Cucuteni-Trypillian culture of Moldova, Romania, and Ukraine
- Roots of a changing early society 7,200-6,700 years ago Jordan and Israel
- Agriculture religion (Paganism) with farming reached Britain between about 7,000 to 6,500 or so years ago and seemingly expressed in things like Western Europe’s Long Barrows
- My Thoughts on Possible Migrations of “R” DNA and Proto-Indo-European?
- “Millet” Spreading from China 7,022 years ago to Europe and related Language may have Spread with it leading to Proto-Indo-European
- Proto-Indo-European (PIE), ancestor of Indo-European languages: DNA, Society, Language, and Mythology
- The Dnieper–Donets culture and Asian varieties of Millet from China to the Black Sea region of Europe by 7,022 years ago
- Kurgan 6,000 years ago/dolmens 7,000 years ago: funeral, ritual, and other?
- 7,020 to 6,020-year-old Proto-Indo-European Homeland of Urheimat or proposed home of their Language and Religion
- Ancient Megaliths: Kurgan, Ziggurat, Pyramid, Menhir, Trilithon, Dolman, Kromlech, and Kromlech of Trilithons
- The Mytheme of Ancient North Eurasian Sacred-Dog belief and similar motifs are found in Indo-European, Native American, and Siberian comparative mythology
- Elite Power Accumulation: Ancient Trade, Tokens, Writing, Wealth, Merchants, and Priest-Kings
- Sacred Mounds, Mountains, Kurgans, and Pyramids may hold deep connections?
- Between 7,000-5,000 Years ago, rise of unequal hierarchy elite, leading to a “birth of the State” or worship of power, strong new sexism, oppression of non-elites, and the fall of Women’s equal status
- Paganism 7,000-5,000 years old: related to “Anarchism and Socialism” (Capitalism) (World War 0) Elite & their slaves
- Hell and Underworld mythologies starting maybe as far back as 7,000 to 5,000 years ago with the Proto-Indo-Europeans?
- The First Expression of the Male God around 7,000 years ago?
- White (light complexion skin) Bigotry and Sexism started 7,000 years ago?
- Around 7,000-year-old Shared Idea of the Divine Bird (Tutelary and/or Trickster spirit/deity), “Ritual” Motif
- Nekhbet an Ancient Egyptian Vulture Goddess and Tutelary Deity
- 6,720 to 4,920 years old Ritualistic Hongshan Culture of Inner Mongolia with 5,000-year-old Pyramid Mounds and Temples
- First proto-king in the Balkans, Varna culture around 6,500 years ago?
- 6,500–5,800 years ago in Israel Late Chalcolithic (Copper Age) Period in the Southern Levant Seems to Express Northern Levant Migrations, Cultural and Religious Transfer
- KING OF BEASTS: Master of Animals “Ritual” Motif, around 6,000 years old or older…
- Around 6000-year-old Shared Idea of the Solid Wheel & the Spoked Wheel-Shaped Ritual Motif
- “The Ghassulian Star,” a mysterious 6,000-year-old mural from Jordan; a Proto-Star of Ishtar, Star of Inanna or Star of Venus?
- Religious/Ritual Ideas, including goddesses and gods as well as ritual mounds or pyramids from Northeastern Asia at least 6,000 years old, seemingly filtering to Iran, Iraq, the Mediterranean, Europe, Egypt, and the Americas?
- Maykop (5,720–5,020 years ago) Caucasus region Bronze Age culture-related to Copper Age farmers from the south, influenced by the Ubaid period and Leyla-Tepe culture, as well as influencing the Kura-Araxes culture
- 5-600-year-old Tomb, Mummy, and First Bearded Male Figurine in a Grave
- Kura-Araxes Cultural 5,520 to 4,470 years old DNA traces to the Canaanites, Arabs, and Jews
- Minoan/Cretan (Keftiu) Civilization and Religion around 5,520 to 3,120 years ago
- Evolution Of Science at least by 5,500 years ago
- 5,500 Years old birth of the State, the rise of Hierarchy, and the fall of Women’s status
- “Jiroft culture” 5,100 – 4,200 years ago and the History of Iran
- Stonehenge: Paganistic Burial and Astrological Ritual Complex, England (5,100-3,600 years ago)
- Around 5,000-year-old Shared Idea of the “Tree of Life” Ritual Motif
- Complex rituals for elite, seen from China to Egypt, at least by 5,000 years ago
- Around 5,000 years ago: “Birth of the State” where Religion gets Military Power and Influence
- The Center of the World “Axis Mundi” and/or “Sacred Mountains” Mythology Could Relate to the Altai Mountains, Heart of the Steppe
- Progressed organized religion starts, an approximately 5,000-year-old belief system
- China’s Civilization between 5,000-3,000 years ago, was a time of war and class struggle, violent transition from free clans to a Slave or Elite society
- Origin of Logics is Naturalistic Observation at least by around 5,000 years ago.
- Paganism 5,000 years old: progressed organized religion and the state: related to “Anarchism and Socialism” (Kings and the Rise of the State)
- Ziggurats (multi-platform temples: 4,900 years old) to Pyramids (multi-platform tombs: 4,700 years old)
- Did a 4,520–4,420-year-old Volcano In Turkey Inspire the Bible God?
- Finland’s Horned Shaman and Pre-Horned-God at least 4,500 years ago?
- 4,000-year-Old Dolmens in Israel: A Connected Dolmen Religious Phenomenon?
- Creation myths: From chaos, Ex nihilo, Earth-diver, Emergence, World egg, and World parent
- Bronze Age “Ritual” connections of the Bell Beaker culture with the Corded Ware/Single Grave culture, which were related to the Yamnaya culture and Proto-Indo-European Languages/Religions
- Low Gods (Earth/ Tutelary deity), High Gods (Sky/Supreme deity), and Moralistic Gods (Deity enforcement/divine order)
- The exchange of people, ideas, and material-culture including, to me, the new god (Sky Father) and goddess (Earth Mother) religion between the Cucuteni-Trypillians and others which is then spread far and wide
- Koryaks: Indigenous People of the Russian Far East and Big Raven myths also found in Tlingit, Haida, Tsimshian, and other Indigenous People of North America
- 42 Principles Of Maat (Egyptian Goddess of the justice) around 4,400 years ago, 2000 Years Before Ten Commandments
- “Happy Easter” Well Happy Eostre/Ishter
- 4,320-3,820 years old “Shimao” (North China) site with Totemistic-Shamanistic Paganism and a Stepped Pyramid
- 4,250 to 3,400 Year old Stonehenge from Russia: Arkaim?
- 4,100-year-old beaker with medicinal & flowering plants in a grave of a woman in Scotland
- Early European Farmer ancestry, Kelif el Boroud people with the Cardial Ware culture, and the Bell Beaker culture Paganists too, spread into North Africa, then to the Canary Islands off West Africa
- Flood Accounts: Gilgamesh epic (4,100 years ago) Noah in Genesis (2,600 years ago)
- Paganism 4,000 years old: related to “Anarchism and Socialism” (First Moralistic gods, then the Origin time of Monotheism)
- When was the beginning: TIMELINE OF CURRENT RELIGIONS, which start around 4,000 years ago.
- Early Religions Thought to Express Proto-Monotheistic Systems around 4,000 years ago
- Kultepe? An archaeological site with a 4,000 years old women’s rights document.
- Single God Religions (Monotheism) = “Man-o-theism” started around 4,000 years ago with the Great Sky Spirit/God Tiān (天)?
- Confucianism’s Tiān (Shangdi god 4,000 years old): Supernaturalism, Pantheism or Theism?
- Yes, Your Male God is Ridiculous
- Mythology, a Lunar Deity is a Goddess or God of the Moon
- Sacred Land, Hills, and Mountains: Sami Mythology (Paganistic Shamanism)
- Horse Worship/Sacrifice: mythical union of Ruling Elite/Kingship and the Horse
- The Amorite/Amurru people’s God Amurru “Lord of the Steppe”, relates to the Origins of the Bible God?
- Bronze Age Exotic Trade Routes Spread Quite Far as well as Spread Religious Ideas with Them
- Sami and the Northern Indigenous Peoples Landscape, Language, and its Connection to Religion
- Prototype of Ancient Analemmatic Sundials around 3,900-3,150 years ago and a Possible Solar Connection to gods?
- Judaism is around 3,450 or 3,250 years old. (“Paleo-Hebrew” 3,000 years ago and Torah 2,500 years ago)
- The Weakening of Ancient Trade and the Strengthening of Religions around 3000 years ago?
- Are you aware that there are religions that worship women gods, explain now religion tears women down?
- Animistic, Totemistic, and Paganistic Superstition Origins of bible god and the bible’s Religion.
- Myths and Folklore: “Trickster gods and goddesses”
- Jews, Judaism, and the Origins of Some of its Ideas
- An Old Branch of Religion Still Giving Fruit: Sacred Trees
- Dating the BIBLE: naming names and telling times (written less than 3,000 years ago, provable to 2,200 years ago)
- Did a Volcano Inspire the bible god?
- Dené–Yeniseian language, Old Copper Complex, and Pre-Columbian Mound Builders?
- No “dinosaurs and humans didn’t exist together just because some think they are in the bible itself”
- Sacred Shit and Sacred Animals?
- Everyone Killed in the Bible Flood? “Nephilim” (giants)?
- Hey, Damien dude, I have a question for you regarding “the bible” Exodus.
- Archaeology Disproves the Bible
- Bible Battle, Just More, Bible Babble
- The Jericho Conquest lie?
- Canaanites and Israelites?
- Accurate Account on how did Christianity Began?
- Let’s talk about Christianity.
- So the 10 commandments isn’t anything to go by either right?
- Misinformed christian
- Debunking Jesus?
- Paulism vs Jesus
- Ok, you seem confused so let’s talk about Buddhism.
- Unacknowledged Buddhism: Gods, Savior, Demons, Rebirth, Heavens, Hells, and Terrorism
- His Foolishness The Dalai Lama
- Yin and Yang is sexist with an ORIGIN around 2,300 years ago?
- I Believe Archaeology, not Myths & Why Not, as the Religious Myths Already Violate Reason!
- Archaeological, Scientific, & Philosophic evidence shows the god myth is man-made nonsense.
- Aquatic Ape Theory/Hypothesis? As Always, Just Pseudoscience.
- Ancient Aliens Conspiracy Theorists are Pseudohistorians
- The Pseudohistoric and Pseudoscientific claims about “Bakoni Ruins” of South Africa
- Why do people think Religion is much more than supernaturalism and superstitionism?
- Religion is an Evolved Product
- Was the Value of Ancient Women Different?
- 1000 to 1100 CE, human sacrifice Cahokia Mounds a pre-Columbian Native American site
- Feminist atheists as far back as the 1800s?
- Promoting Religion as Real is Mentally Harmful to a Flourishing Humanity
- Screw All Religions and Their Toxic lies, they are all fraud
- Forget Religions’ Unfounded Myths, I Have Substantiated “Archaeology Facts.”
- Religion Dispersal throughout the World
- I Hate Religion Just as I Hate all Pseudoscience
- Exposing Scientology, Eckankar, Wicca and Other Nonsense?
- Main deity or religious belief systems
- Quit Trying to Invent Your God From the Scraps of Science.
- Archaeological, Scientific, & Philosophic evidence shows the god myth is man-made nonsense.
- Ancient Alien Conspiracy Theorists: Misunderstanding, Rhetoric, Misinformation, Fabrications, and Lies
- Misinformation, Distortion, and Pseudoscience in Talking with a Christian Creationist
- Judging the Lack of Goodness in Gods, Even the Norse God Odin
- Challenging the Belief in God-like Aliens and Gods in General
- A Challenge to Christian use of Torture Devices?
- Yes, Hinduism is a Religion
- Trump is One of the Most Reactionary Forces of Far-right Christian Extremism
- Was the Bull Head a Symbol of God? Yes!
- Primate Death Rituals
- Christian – “God and Christianity are objectively true”
- Australopithecus afarensis Death Ritual?
- You Claim Global Warming is a Hoax?
- Doubter of Science and Defamer of Atheists?
- I think that sounds like the Bible?
- History of the Antifa (“anti-fascist”) Movements
- Indianapolis Anti-Blasphemy Laws #Free Soheil Rally
- Damien, you repeat the golden rule in so many forms then you say religion is dogmatic?
- Science is a Trustable Methodology whereas Faith is not Trustable at all!
- Was I ever a believer, before I was an atheist?
- Atheists rise in reason
- Mistrust of science?
- Open to Talking About the Definition of ‘God’? But first, we address Faith.
- ‘United Monarchy’ full of splendor and power – Saul, David, and Solomon? Most likely not.
- Is there EXODUS ARCHAEOLOGY? The short answer is “no.”
- Lacking Proof of Bigfoots, Unicorns, and Gods is Just a Lack of Research?
- Religion and Politics: Faith Beliefs vs. Rational Thinking
- Hammer of Truth that lying pig RELIGION: challenged by an archaeologist
- “The Hammer of Truth” -ontology question- What do You Mean by That?
- Navigation of a bad argument: Ad Hominem vs. Attack
- Why is it Often Claimed that Gods have a Gender?
- Why are basically all monotheistic religions ones that have a male god?
- Shifting through the Claims in support of Faith
- Dear Mr. AtHope, The 20th Century is an Indictment of Secularism and a Failed Atheist Century
- An Understanding of the Worldwide Statistics and Dynamics of Terrorist Incidents and Suicide Attacks
- Intoxication and Evolution? Addressing and Assessing the “Stoned Ape” or “Drunken Monkey” Theories as Catalysts in Human Evolution
- Sacred Menstrual cloth? Inanna’s knot, Isis knot, and maybe Ma’at’s feather?
- Damien, why don’t the Hebrews accept the bible stories?
- Dealing with a Troll and Arguing Over Word Meaning
- Knowledge without Belief? Justified beliefs or disbeliefs worthy of Knowledge?
- Afrocentrism and African Religions
- Crecganford @crecganford offers history & stories of the people, places, gods, & culture
- Empiricism-Denier?
I am not an academic. I am a revolutionary that teaches in public, in places like social media, and in the streets. I am not a leader by some title given but from my commanding leadership style of simply to start teaching everywhere to everyone, all manner of positive education.



To me, Animism starts in Southern Africa, then to West Europe, and becomes Totemism. Another split goes near the Russia and Siberia border becoming Shamanism, which heads into Central Europe meeting up with Totemism, which also had moved there, mixing the two which then heads to Lake Baikal in Siberia. From there this Shamanism-Totemism heads to Turkey where it becomes Paganism.



-LAPTOP-KIB0FGT7")

Not all “Religions” or “Religious Persuasions” have a god(s) but
All can be said to believe in some imaginary beings or imaginary things like spirits, afterlives, etc.

{kind=link}
Paganism 12,000-4,000 years old
12,000-7,000 years old: related to (Pre-Capitalism)
7,000-5,000 years old: related to (Capitalism) (World War 0) Elite and their slaves!
5,000 years old: related to (Kings and the Rise of the State)
4,000 years old: related to (First Moralistic gods, then the Origin time of Monotheism)

ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref
Low Gods “Earth” or Tutelary deity and High Gods “Sky” or Supreme deity
“An Earth goddess is a deification of the Earth. Earth goddesses are often associated with the “chthonic” deities of the underworld. Ki and Ninhursag are Mesopotamian earth goddesses. In Greek mythology, the Earth is personified as Gaia, corresponding to Roman Terra, Indic Prithvi/Bhūmi, etc. traced to an “Earth Mother” complementary to the “Sky Father” in Proto-Indo-European religion. Egyptian mythology exceptionally has a sky goddess and an Earth god.” ref
“A mother goddess is a goddess who represents or is a personification of nature, motherhood, fertility, creation, destruction or who embodies the bounty of the Earth. When equated with the Earth or the natural world, such goddesses are sometimes referred to as Mother Earth or as the Earth Mother. In some religious traditions or movements, Heavenly Mother (also referred to as Mother in Heaven or Sky Mother) is the wife or feminine counterpart of the Sky father or God the Father.” ref
“Any masculine sky god is often also king of the gods, taking the position of patriarch within a pantheon. Such king gods are collectively categorized as “sky father” deities, with a polarity between sky and earth often being expressed by pairing a “sky father” god with an “earth mother” goddess (pairings of a sky mother with an earth father are less frequent). A main sky goddess is often the queen of the gods and may be an air/sky goddess in her own right, though she usually has other functions as well with “sky” not being her main. In antiquity, several sky goddesses in ancient Egypt, Mesopotamia, and the Near East were called Queen of Heaven. Neopagans often apply it with impunity to sky goddesses from other regions who were never associated with the term historically. The sky often has important religious significance. Many religions, both polytheistic and monotheistic, have deities associated with the sky.” ref
“In comparative mythology, sky father is a term for a recurring concept in polytheistic religions of a sky god who is addressed as a “father”, often the father of a pantheon and is often either a reigning or former King of the Gods. The concept of “sky father” may also be taken to include Sun gods with similar characteristics, such as Ra. The concept is complementary to an “earth mother“. “Sky Father” is a direct translation of the Vedic Dyaus Pita, etymologically descended from the same Proto-Indo-European deity name as the Greek Zeûs Pater and Roman Jupiter and Germanic Týr, Tir or Tiwaz, all of which are reflexes of the same Proto-Indo-European deity’s name, *Dyēus Ph₂tḗr. While there are numerous parallels adduced from outside of Indo-European mythology, there are exceptions (e.g. In Egyptian mythology, Nut is the sky mother and Geb is the earth father).” ref
Tutelary deity
“A tutelary (also tutelar) is a deity or spirit who is a guardian, patron, or protector of a particular place, geographic feature, person, lineage, nation, culture, or occupation. The etymology of “tutelary” expresses the concept of safety and thus of guardianship. In late Greek and Roman religion, one type of tutelary deity, the genius, functions as the personal deity or daimon of an individual from birth to death. Another form of personal tutelary spirit is the familiar spirit of European folklore.” ref
“A tutelary (also tutelar) in Korean shamanism, jangseung and sotdae were placed at the edge of villages to frighten off demons. They were also worshiped as deities. Seonangshin is the patron deity of the village in Korean tradition and was believed to embody the Seonangdang. In Philippine animism, Diwata or Lambana are deities or spirits that inhabit sacred places like mountains and mounds and serve as guardians. Such as: Maria Makiling is the deity who guards Mt. Makiling and Maria Cacao and Maria Sinukuan. In Shinto, the spirits, or kami, which give life to human bodies come from nature and return to it after death. Ancestors are therefore themselves tutelaries to be worshiped. And similarly, Native American beliefs such as Tonás, tutelary animal spirit among the Zapotec and Totems, familial or clan spirits among the Ojibwe, can be animals.” ref
“A tutelary (also tutelar) in Austronesian beliefs such as: Atua (gods and spirits of the Polynesian peoples such as the Māori or the Hawaiians), Hanitu (Bunun of Taiwan‘s term for spirit), Hyang (Kawi, Sundanese, Javanese, and Balinese Supreme Being, in ancient Java and Bali mythology and this spiritual entity, can be either divine or ancestral), Kaitiaki (New Zealand Māori term used for the concept of guardianship, for the sky, the sea, and the land), Kawas (mythology) (divided into 6 groups: gods, ancestors, souls of the living, spirits of living things, spirits of lifeless objects, and ghosts), Tiki (Māori mythology, Tiki is the first man created by either Tūmatauenga or Tāne and represents deified ancestors found in most Polynesian cultures). ” ref, ref, ref, ref, ref, ref, ref
Mesopotamian Tutelary Deities can be seen as ones related to City-States
“Historical city-states included Sumerian cities such as Uruk and Ur; Ancient Egyptian city-states, such as Thebes and Memphis; the Phoenician cities (such as Tyre and Sidon); the five Philistine city-states; the Berber city-states of the Garamantes; the city-states of ancient Greece (the poleis such as Athens, Sparta, Thebes, and Corinth); the Roman Republic (which grew from a city-state into a vast empire); the Italian city-states from the Middle Ages to the early modern period, such as Florence, Siena, Ferrara, Milan (which as they grew in power began to dominate neighboring cities) and Genoa and Venice, which became powerful thalassocracies; the Mayan and other cultures of pre-Columbian Mesoamerica (including cities such as Chichen Itza, Tikal, Copán and Monte Albán); the central Asian cities along the Silk Road; the city-states of the Swahili coast; Ragusa; states of the medieval Russian lands such as Novgorod and Pskov; and many others.” ref
“The Uruk period (ca. 4000 to 3100 BCE; also known as Protoliterate period) of Mesopotamia, named after the Sumerian city of Uruk, this period saw the emergence of urban life in Mesopotamia and the Sumerian civilization. City-States like Uruk and others had a patron tutelary City Deity along with a Priest-King.” ref
“Chinese folk religion, both past, and present, includes myriad tutelary deities. Exceptional individuals, highly cultivated sages, and prominent ancestors can be deified and honored after death. Lord Guan is the patron of military personnel and police, while Mazu is the patron of fishermen and sailors. Such as Tu Di Gong (Earth Deity) is the tutelary deity of a locality, and each individual locality has its own Earth Deity and Cheng Huang Gong (City God) is the guardian deity of an individual city, worshipped by local officials and locals since imperial times.” ref
“A tutelary (also tutelar) in Hinduism, personal tutelary deities are known as ishta-devata, while family tutelary deities are known as Kuladevata. Gramadevata are guardian deities of villages. Devas can also be seen as tutelary. Shiva is the patron of yogis and renunciants. City goddesses include: Mumbadevi (Mumbai), Sachchika (Osian); Kuladevis include: Ambika (Porwad), and Mahalakshmi. In NorthEast India Meitei mythology and religion (Sanamahism) of Manipur, there are various types of tutelary deities, among which Lam Lais are the most predominant ones. Tibetan Buddhism has Yidam as a tutelary deity. Dakini is the patron of those who seek knowledge.” ref
“A tutelary (also tutelar) The Greeks also thought deities guarded specific places: for instance, Athena was the patron goddess of the city of Athens. Socrates spoke of hearing the voice of his personal spirit or daimonion:
You have often heard me speak of an oracle or sign which comes to me … . This sign I have had ever since I was a child. The sign is a voice which comes to me and always forbids me to do something which I am going to do, but never commands me to do anything, and this is what stands in the way of my being a politician.” ref
“Tutelary deities who guard and preserve a place or a person are fundamental to ancient Roman religion. The tutelary deity of a man was his Genius, that of a woman her Juno. In the Imperial era, the Genius of the Emperor was a focus of Imperial cult. An emperor might also adopt a major deity as his personal patron or tutelary, as Augustus did Apollo. Precedents for claiming the personal protection of a deity were established in the Republican era, when for instance the Roman dictator Sulla advertised the goddess Victory as his tutelary by holding public games (ludi) in her honor.” ref
“Each town or city had one or more tutelary deities, whose protection was considered particularly vital in time of war and siege. Rome itself was protected by a goddess whose name was to be kept ritually secret on pain of death (for a supposed case, see Quintus Valerius Soranus). The Capitoline Triad of Juno, Jupiter, and Minerva were also tutelaries of Rome. The Italic towns had their own tutelary deities. Juno often had this function, as at the Latin town of Lanuvium and the Etruscan city of Veii, and was often housed in an especially grand temple on the arx (citadel) or other prominent or central location. The tutelary deity of Praeneste was Fortuna, whose oracle was renowned.” ref
“The Roman ritual of evocatio was premised on the belief that a town could be made vulnerable to military defeat if the power of its tutelary deity were diverted outside the city, perhaps by the offer of superior cult at Rome. The depiction of some goddesses such as the Magna Mater (Great Mother, or Cybele) as “tower-crowned” represents their capacity to preserve the city. A town in the provinces might adopt a deity from within the Roman religious sphere to serve as its guardian, or syncretize its own tutelary with such; for instance, a community within the civitas of the Remi in Gaul adopted Apollo as its tutelary, and at the capital of the Remi (present-day Rheims), the tutelary was Mars Camulus.” ref
Household deity (a kind of or related to a Tutelary deity)
“A household deity is a deity or spirit that protects the home, looking after the entire household or certain key members. It has been a common belief in paganism as well as in folklore across many parts of the world. Household deities fit into two types; firstly, a specific deity – typically a goddess – often referred to as a hearth goddess or domestic goddess who is associated with the home and hearth, such as the ancient Greek Hestia.” ref
“The second type of household deities are those that are not one singular deity, but a type, or species of animistic deity, who usually have lesser powers than major deities. This type was common in the religions of antiquity, such as the Lares of ancient Roman religion, the Gashin of Korean shamanism, and Cofgodas of Anglo-Saxon paganism. These survived Christianisation as fairy-like creatures existing in folklore, such as the Anglo-Scottish Brownie and Slavic Domovoy.” ref
“Household deities were usually worshipped not in temples but in the home, where they would be represented by small idols (such as the teraphim of the Bible, often translated as “household gods” in Genesis 31:19 for example), amulets, paintings, or reliefs. They could also be found on domestic objects, such as cosmetic articles in the case of Tawaret. The more prosperous houses might have a small shrine to the household god(s); the lararium served this purpose in the case of the Romans. The gods would be treated as members of the family and invited to join in meals, or be given offerings of food and drink.” ref
“In many religions, both ancient and modern, a god would preside over the home. Certain species, or types, of household deities, existed. An example of this was the Roman Lares. Many European cultures retained house spirits into the modern period. Some examples of these include:
- Brownie (Scotland and England) or Hob (England) / Kobold (Germany) / Goblin / Hobgoblin
- Domovoy (Slavic)
- Nisse (Norwegian or Danish) / Tomte (Swedish) / Tonttu (Finnish)
- Húsvættir (Norse)” ref
“Although the cosmic status of household deities was not as lofty as that of the Twelve Olympians or the Aesir, they were also jealous of their dignity and also had to be appeased with shrines and offerings, however humble. Because of their immediacy they had arguably more influence on the day-to-day affairs of men than the remote gods did. Vestiges of their worship persisted long after Christianity and other major religions extirpated nearly every trace of the major pagan pantheons. Elements of the practice can be seen even today, with Christian accretions, where statues to various saints (such as St. Francis) protect gardens and grottos. Even the gargoyles found on older churches, could be viewed as guardians partitioning a sacred space.” ref
“For centuries, Christianity fought a mop-up war against these lingering minor pagan deities, but they proved tenacious. For example, Martin Luther‘s Tischreden have numerous – quite serious – references to dealing with kobolds. Eventually, rationalism and the Industrial Revolution threatened to erase most of these minor deities, until the advent of romantic nationalism rehabilitated them and embellished them into objects of literary curiosity in the 19th century. Since the 20th century this literature has been mined for characters for role-playing games, video games, and other fantasy personae, not infrequently invested with invented traits and hierarchies somewhat different from their mythological and folkloric roots.” ref
“In contradistinction to both Herbert Spencer and Edward Burnett Tylor, who defended theories of animistic origins of ancestor worship, Émile Durkheim saw its origin in totemism. In reality, this distinction is somewhat academic, since totemism may be regarded as a particularized manifestation of animism, and something of a synthesis of the two positions was attempted by Sigmund Freud. In Freud’s Totem and Taboo, both totem and taboo are outward expressions or manifestations of the same psychological tendency, a concept which is complementary to, or which rather reconciles, the apparent conflict. Freud preferred to emphasize the psychoanalytic implications of the reification of metaphysical forces, but with particular emphasis on its familial nature. This emphasis underscores, rather than weakens, the ancestral component.” ref
“William Edward Hearn, a noted classicist, and jurist, traced the origin of domestic deities from the earliest stages as an expression of animism, a belief system thought to have existed also in the neolithic, and the forerunner of Indo-European religion. In his analysis of the Indo-European household, in Chapter II “The House Spirit”, Section 1, he states:
The belief which guided the conduct of our forefathers was … the spirit rule of dead ancestors.” ref
“In Section 2 he proceeds to elaborate:
It is thus certain that the worship of deceased ancestors is a vera causa, and not a mere hypothesis. …
In the other European nations, the Slavs, the Teutons, and the Kelts, the House Spirit appears with no less distinctness. … [T]he existence of that worship does not admit of doubt. … The House Spirits had a multitude of other names which it is needless here to enumerate, but all of which are more or less expressive of their friendly relations with man. … In [England] … [h]e is the Brownie. … In Scotland this same Brownie is well known. He is usually described as attached to particular families, with whom he has been known to reside for centuries, threshing the corn, cleaning the house, and performing similar household tasks. His favorite gratification was milk and honey.” ref

ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref
“These ideas are my speculations from the evidence.”
I am still researching the “god‘s origins” all over the world. So you know, it is very complicated but I am smart and willing to look, DEEP, if necessary, which going very deep does seem to be needed here, when trying to actually understand the evolution of gods and goddesses. I am sure of a few things and less sure of others, but even in stuff I am not fully grasping I still am slowly figuring it out, to explain it to others. But as I research more I am understanding things a little better, though I am still working on understanding it all or something close and thus always figuring out more.
Sky Father/Sky God?
“Egyptian: (Nut) Sky Mother and (Geb) Earth Father” (Egypt is different but similar)
Turkic/Mongolic: (Tengri/Tenger Etseg) Sky Father and (Eje/Gazar Eej) Earth Mother *Transeurasian*
Hawaiian: (Wākea) Sky Father and (Papahānaumoku) Earth Mother *Austronesian*
New Zealand/ Māori: (Ranginui) Sky Father and (Papatūānuku) Earth Mother *Austronesian*
Proto-Indo-European: (Dyḗus/Dyḗus ph₂tḗr) Sky Father and (Dʰéǵʰōm/Pleth₂wih₁) Earth Mother
Indo-Aryan: (Dyaus Pita) Sky Father and (Prithvi Mata) Earth Mother *Indo-European*
Italic: (Jupiter) Sky Father and (Juno) Sky Mother *Indo-European*
Etruscan: (Tinia) Sky Father and (Uni) Sky Mother *Tyrsenian/Italy Pre–Indo-European*
Hellenic/Greek: (Zeus) Sky Father and (Hera) Sky Mother who started as an “Earth Goddess” *Indo-European*
Nordic: (Dagr) Sky Father and (Nótt) Sky Mother *Indo-European*
Slavic: (Perun) Sky Father and (Mokosh) Earth Mother *Indo-European*
Illyrian: (Deipaturos) Sky Father and (Messapic Damatura’s “earth-mother” maybe) Earth Mother *Indo-European*
Albanian: (Zojz) Sky Father and (?) *Indo-European*
Baltic: (Perkūnas) Sky Father and (Saulė) Sky Mother *Indo-European*
Germanic: (Týr) Sky Father and (?) *Indo-European*
Colombian-Muisca: (Bochica) Sky Father and (Huythaca) Sky Mother *Chibchan*
Aztec: (Quetzalcoatl) Sky Father and (Xochiquetzal) Sky Mother *Uto-Aztecan*
Incan: (Viracocha) Sky Father and (Mama Runtucaya) Sky Mother *Quechuan*
China: (Tian/Shangdi) Sky Father and (Dì) Earth Mother *Sino-Tibetan*
Sumerian, Assyrian and Babylonian: (An/Anu) Sky Father and (Ki) Earth Mother
Finnish: (Ukko) Sky Father and (Akka) Earth Mother *Finno-Ugric*
Sami: (Horagalles) Sky Father and (Ravdna) Earth Mother *Finno-Ugric*
Puebloan-Zuni: (Ápoyan Ta’chu) Sky Father and (Áwitelin Tsíta) Earth Mother
Puebloan-Hopi: (Tawa) Sky Father and (Kokyangwuti/Spider Woman/Grandmother) Earth Mother *Uto-Aztecan*
Puebloan-Navajo: (Tsohanoai) Sky Father and (Estsanatlehi) Earth Mother *Na-Dene*
ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref, ref

Hinduism around 3,700 to 3,500 years old. ref
Judaism around 3,450 or 3,250 years old. (The first writing in the bible was “Paleo-Hebrew” dated to around 3,000 years ago Khirbet Qeiyafa is the site of an ancient fortress city overlooking the Elah Valley. And many believe the religious Jewish texts were completed around 2,500) ref, ref
Judaism is around 3,450 or 3,250 years old. (“Paleo-Hebrew” 3,000 years ago and Torah 2,500 years ago)
“Judaism is an Abrahamic, its roots as an organized religion in the Middle East during the Bronze Age. Some scholars argue that modern Judaism evolved from Yahwism, the religion of ancient Israel and Judah, by the late 6th century BCE, and is thus considered to be one of the oldest monotheistic religions.” ref
“Yahwism is the name given by modern scholars to the religion of ancient Israel, essentially polytheistic, with a plethora of gods and goddesses. Heading the pantheon was Yahweh, the national god of the Israelite kingdoms of Israel and Judah, with his consort, the goddess Asherah; below them were second-tier gods and goddesses such as Baal, Shamash, Yarikh, Mot, and Astarte, all of whom had their own priests and prophets and numbered royalty among their devotees, and a third and fourth tier of minor divine beings, including the mal’ak, the messengers of the higher gods, who in later times became the angels of Judaism, Christianity and Islam. Yahweh, however, was not the ‘original’ god of Israel “Isra-El”; it is El, the head of the Canaanite pantheon, whose name forms the basis of the name “Israel”, and none of the Old Testament patriarchs, the tribes of Israel, the Judges, or the earliest monarchs, have a Yahwistic theophoric name (i.e., one incorporating the name of Yahweh).” ref
“El is a Northwest Semitic word meaning “god” or “deity“, or referring (as a proper name) to any one of multiple major ancient Near Eastern deities. A rarer form, ‘ila, represents the predicate form in Old Akkadian and in Amorite. The word is derived from the Proto-Semitic *ʔil-, meaning “god”. Specific deities known as ‘El or ‘Il include the supreme god of the ancient Canaanite religion and the supreme god of East Semitic speakers in Mesopotamia’s Early Dynastic Period. ʼĒl is listed at the head of many pantheons. In some Canaanite and Ugaritic sources, ʼĒl played a role as father of the gods, of creation, or both. For example, in the Ugaritic texts, ʾil mlk is understood to mean “ʼĒl the King” but ʾil hd as “the god Hadad“. The Semitic root ʾlh (Arabic ʾilāh, Aramaic ʾAlāh, ʾElāh, Hebrew ʾelōah) may be ʾl with a parasitic h, and ʾl may be an abbreviated form of ʾlh. In Ugaritic the plural form meaning “gods” is ʾilhm, equivalent to Hebrew ʾelōhîm “powers”. In the Hebrew texts this word is interpreted as being semantically singular for “god” by biblical commentators. However the documentary hypothesis for the Old Testament (corresponds to the Jewish Torah) developed originally in the 1870s, identifies these that different authors – the Jahwist, Elohist, Deuteronomist, and the Priestly source – were responsible for editing stories from a polytheistic religion into those of a monotheistic religion. Inconsistencies that arise between monotheism and polytheism in the texts are reflective of this hypothesis.” ref
Jainism around 2,599 – 2,527 years old. ref
Confucianism around 2,600 – 2,551 years old. ref
Buddhism around 2,563/2,480 – 2,483/2,400 years old. ref
Christianity around 2,o00 years old. ref
Shinto around 1,305 years old. ref
Islam around 1407–1385 years old. ref

Knowledge to Ponder:
Stars/Astrology:
- Possibly, around 30,000 years ago (in simpler form) to 6,000 years ago, Stars/Astrology are connected to Ancestors, Spirit Animals, and Deities.
- The star also seems to be a possible proto-star for Star of Ishtar, Star of Inanna, or Star of Venus.
- Around 7,000 to 6,000 years ago, Star Constellations/Astrology have connections to the “Kurgan phenomenon” of below-ground “mound” stone/wood burial structures and “Dolmen phenomenon” of above-ground stone burial structures.
- Around 6,500–5,800 years ago, The Northern Levant migrations into Jordon and Israel in the Southern Levant brought new cultural and religious transfer from Turkey and Iran.
- “The Ghassulian Star,” a mysterious 6,000-year-old mural from Jordan may have connections to the European paganstic kurgan/dolmens phenomenon.
“Astrology is a range of divinatory practices, recognized as pseudoscientific since the 18th century, that claim to discern information about human affairs and terrestrial events by studying the apparent positions of celestial objects. Different cultures have employed forms of astrology since at least the 2nd millennium BCE, these practices having originated in calendrical systems used to predict seasonal shifts and to interpret celestial cycles as signs of divine communications. Most, if not all, cultures have attached importance to what they observed in the sky, and some—such as the Hindus, Chinese, and the Maya—developed elaborate systems for predicting terrestrial events from celestial observations. Western astrology, one of the oldest astrological systems still in use, can trace its roots to 19th–17th century BCE Mesopotamia, from where it spread to Ancient Greece, Rome, the Islamicate world and eventually Central and Western Europe. Contemporary Western astrology is often associated with systems of horoscopes that purport to explain aspects of a person’s personality and predict significant events in their lives based on the positions of celestial objects; the majority of professional astrologers rely on such systems.” ref
Around 5,500 years ago, Science evolves, The first evidence of science was 5,500 years ago and was demonstrated by a body of empirical, theoretical, and practical knowledge about the natural world. ref
Around 5,000 years ago, Origin of Logics is a Naturalistic Observation (principles of valid reasoning, inference, & demonstration) ref
Around 4,150 to 4,000 years ago: The earliest surviving versions of the Sumerian Epic of Gilgamesh, which was originally titled “He who Saw the Deep” (Sha naqba īmuru) or “Surpassing All Other Kings” (Shūtur eli sharrī) were written. ref
Hinduism:
- 3,700 years ago or so, the oldest of the Hindu Vedas (scriptures), the Rig Veda was composed.
- 3,500 years ago or so, the Vedic Age began in India after the collapse of the Indus Valley Civilization.
Judaism:
- around 3,000 years ago, the first writing in the bible was “Paleo-Hebrew”
- around 2,500 years ago, many believe the religious Jewish texts were completed
Myths: The bible inspired religion is not just one religion or one myth but a grouping of several religions and myths
- Around 3,450 or 3,250 years ago, according to legend, is the traditionally accepted period in which the Israelite lawgiver, Moses, provided the Ten Commandments.
- Around 2,500 to 2,400 years ago, a collection of ancient religious writings by the Israelites based primarily upon the Hebrew Bible, Tanakh, or Old Testament is the first part of Christianity’s bible.
- Around 2,400 years ago, the most accepted hypothesis is that the canon was formed in stages, first the Pentateuch (Torah).
- Around 2,140 to 2,116 years ago, the Prophets was written during the Hasmonean dynasty, and finally the remaining books.
- Christians traditionally divide the Old Testament into four sections:
- The first five books or Pentateuch (Torah).
- The proposed history books telling the history of the Israelites from their conquest of Canaan to their defeat and exile in Babylon.
- The poetic and proposed “Wisdom books” dealing, in various forms, with questions of good and evil in the world.
- The books of the biblical prophets, warning of the consequences of turning away from God:
- Henotheism:
- Exodus 20:23 “You shall not make other gods besides Me (not saying there are no other gods just not to worship them); gods of silver or gods of gold, you shall not make for yourselves.”
- Polytheism:
- Judges 10:6 “Then the sons of Israel again did evil in the sight of the LORD, served the Baals and the Ashtaroth, the gods of Aram, the gods of Sidon, the gods of Moab, the gods of the sons of Ammon, and the gods of the Philistines; thus they forsook the LORD and did not serve Him.”
- 1 Corinthians 8:5 “For even if there are so-called gods whether in heaven or on earth, as indeed there are many gods and many lords.”
- Monotheism:
- Isaiah 43:10 “You are my witnesses,” declares the LORD, “and my servant whom I have chosen, so that you may know and believe me and understand that I am he. Before me no god was formed, nor will there be one after me.
Around 2,570 to 2,270 Years Ago, there is a confirmation of atheistic doubting as well as atheistic thinking, mainly by Greek philosophers. However, doubting gods is likely as old as the invention of gods and should destroy the thinking that belief in god(s) is the “default belief”. The Greek word is apistos (a “not” and pistos “faithful,”), thus not faithful or faithless because one is unpersuaded and unconvinced by a god(s) claim. Short Definition: unbelieving, unbeliever, or unbelief.

Expressions of Atheistic Thinking:
- Around 2,600 years ago, Ajita Kesakambali, ancient Indian philosopher, who is the first known proponent of Indian materialism. ref
- Around 2,535 to 2,475 years ago, Heraclitus, Greek pre-Socratic philosopher, a native of the Greek city Ephesus, Ionia, on the coast of Anatolia, also known as Asia Minor or modern Turkey. ref
- Around 2,500 to 2,400 years ago, according to The Story of Civilization book series certain African pygmy tribes have no identifiable gods, spirits, or religious beliefs or rituals, and even what burials accrue are without ceremony. ref
- Around 2,490 to 2,430 years ago, Empedocles, Greek pre-Socratic philosopher and a citizen of Agrigentum, a Greek city in Sicily. ref
- Around 2,460 to 2,370 years ago, Democritus, Greek pre-Socratic philosopher considered to be the “father of modern science” possibly had some disbelief amounting to atheism. ref
- Around 2,399 years ago or so, Socrates, a famous Greek philosopher was tried for sinfulness by teaching doubt of state gods. ref
- Around 2,341 to 2,270 years ago, Epicurus, a Greek philosopher known for composing atheistic critics and famously stated, “Is God willing to prevent evil, but not able? Then he is not omnipotent. Is he able, but not willing? Then he is malevolent. Is he both able and willing? Then whence cometh evil? Is he neither able nor willing? Then why call him god?” ref
This last expression by Epicurus, seems to be an expression of Axiological Atheism. To understand and utilize value or actually possess “Value Conscious/Consciousness” to both give a strong moral “axiological” argument (the problem of evil) as well as use it to fortify humanism and positive ethical persuasion of human helping and care responsibilities. Because value-blindness gives rise to sociopathic/psychopathic evil.

“Theists, there has to be a god, as something can not come from nothing.”
Well, thus something (unknown) happened and then there was something. This does not tell us what the something that may have been involved with something coming from nothing. A supposed first cause, thus something (unknown) happened and then there was something is not an open invitation to claim it as known, neither is it justified to call or label such an unknown as anything, especially an unsubstantiated magical thinking belief born of mythology and religious storytelling.
")

While hallucinogens are associated with shamanism, it is alcohol that is associated with paganism.
The Atheist-Humanist-Leftist Revolutionaries Shows in the prehistory series:
Show two: Pre-animism 300,000 years old and animism 100,000 years old: related to “Anarchism and Socialism”
Show tree: Totemism 50,000 years old: related to “Anarchism and Socialism”
Show four: Shamanism 30,000 years old: related to “Anarchism and Socialism”
Show five: Paganism 12,000 years old: related to “Anarchism and Socialism”
Show six: Emergence of hierarchy, sexism, slavery, and the new male god dominance: Paganism 7,000-5,000 years old: related to “Anarchism and Socialism” (Capitalism) (World War 0) Elite and their slaves!
Prehistory: related to “Anarchism and Socialism” the division of labor, power, rights, and recourses: VIDEO
Pre-animism 300,000 years old and animism 100,000 years old: related to “Anarchism and Socialism”: VIDEO
Totemism 50,000 years old: related to “Anarchism and Socialism”: VIDEO
Shamanism 30,000 years old: related to “Anarchism and Socialism”: VIDEO
Paganism 12,000 years old: related to “Anarchism and Socialism” (Pre-Capitalism): VIDEO
Paganism 7,000-5,000 years old: related to “Anarchism and Socialism” (Capitalism) (World War 0) Elite and their slaves: VIEDO
Paganism 5,000 years old: progressed organized religion and the state: related to “Anarchism and Socialism” (Kings and the Rise of the State): VIEDO
Paganism 4,000 years old: related to “Anarchism and Socialism” (First Moralistic gods, then the Origin time of Monotheism): VIEDO
I do not hate simply because I challenge and expose myths or lies any more than others being thought of as loving simply because of the protection and hiding from challenge their favored myths or lies.
The truth is best championed in the sunlight of challenge.
An archaeologist once said to me “Damien religion and culture are very different”
My response, So are you saying that was always that way, such as would you say Native Americans’ cultures are separate from their religions? And do you think it always was the way you believe?
I had said that religion was a cultural product. That is still how I see it and there are other archaeologists that think close to me as well. Gods too are the myths of cultures that did not understand science or the world around them, seeing magic/supernatural everywhere.
I personally think there is a goddess and not enough evidence to support a male god at Çatalhöyük but if there was both a male and female god and goddess then I know the kind of gods they were like Proto-Indo-European mythology.
This series idea was addressed in, Anarchist Teaching as Free Public Education or Free Education in the Public: VIDEO
Our 12 video series: Organized Oppression: Mesopotamian State Force and the Politics of power (9,000-4,000 years ago), is adapted from: The Complete and Concise History of the Sumerians and Early Bronze Age Mesopotamia (7000-2000 BC): https://www.youtube.com/watch?v=szFjxmY7jQA by “History with Cy“
Show #1: Mesopotamian State Force and the Politics of Power (Samarra, Halaf, Ubaid)
Show #2: Mesopotamian State Force and the Politics of Power
Show #3: Mesopotamian State Force and the Politics of Power (Uruk and the First Cities)
Show #4: Mesopotamian State Force and the Politics of Power (First Kings)
Show #5: Mesopotamian State Force and the Politics of Power (Early Dynastic Period)
Show #6: Mesopotamian State Force and the Politics of Power
Show #7: Mesopotamian State Force and the Politics of Power (Sargon and Akkadian Rule)
Show #9: Mesopotamian State Force and the Politics of Power (Gudea of Lagash and Utu-hegal)
Show #12: Mesopotamian State Force and the Politics of Power (Aftermath and Legacy of Sumer)

The “Atheist-Humanist-Leftist Revolutionaries”
Cory Johnston ☭ Ⓐ Atheist Leftist @Skepticallefty & I (Damien Marie AtHope) @AthopeMarie (my YouTube & related blog) are working jointly in atheist, antitheist, antireligionist, antifascist, anarchist, socialist, and humanist endeavors in our videos together, generally, every other Saturday.
Why Does Power Bring Responsibility?
Think, how often is it the powerless that start wars, oppress others, or commit genocide? So, I guess the question is to us all, to ask, how can power not carry responsibility in a humanity concept? I know I see the deep ethical responsibility that if there is power their must be a humanistic responsibility of ethical and empathic stewardship of that power. Will I be brave enough to be kind? Will I possess enough courage to be compassionate? Will my valor reach its height of empathy? I as everyone, earns our justified respect by our actions, that are good, ethical, just, protecting, and kind. Do I have enough self-respect to put my love for humanity’s flushing, over being brought down by some of its bad actors? May we all be the ones doing good actions in the world, to help human flourishing.
I create the world I want to live in, striving for flourishing. Which is not a place but a positive potential involvement and promotion; a life of humanist goal precision. To master oneself, also means mastering positive prosocial behaviors needed for human flourishing. I may have lost a god myth as an atheist, but I am happy to tell you, my friend, it is exactly because of that, leaving the mental terrorizer, god belief, that I truly regained my connected ethical as well as kind humanity.
Cory and I will talk about prehistory and theism, addressing the relevance to atheism, anarchism, and socialism.
At the same time as the rise of the male god, 7,000 years ago, there was also the very time there was the rise of violence, war, and clans to kingdoms, then empires, then states. It is all connected back to 7,000 years ago, and it moved across the world.
Cory Johnston: https://damienmarieathope.com/2021/04/cory-johnston-mind-of-a-skeptical-leftist/?v=32aec8db952d
The Mind of a Skeptical Leftist (YouTube)
Cory Johnston: Mind of a Skeptical Leftist @Skepticallefty
The Mind of a Skeptical Leftist By Cory Johnston: “Promoting critical thinking, social justice, and left-wing politics by covering current events and talking to a variety of people. Cory Johnston has been thoughtfully talking to people and attempting to promote critical thinking, social justice, and left-wing politics.” http://anchor.fm/skepticalleft
Cory needs our support. We rise by helping each other.
Cory Johnston ☭ Ⓐ @Skepticallefty Evidence-based atheist leftist (he/him) Producer, host, and co-host of 4 podcasts @skeptarchy @skpoliticspod and @AthopeMarie
Damien Marie AtHope (“At Hope”) Axiological Atheist, Anti-theist, Anti-religionist, Secular Humanist. Rationalist, Writer, Artist, Poet, Philosopher, Advocate, Activist, Psychology, and Armchair Archaeology/Anthropology/Historian.
Damien is interested in: Freedom, Liberty, Justice, Equality, Ethics, Humanism, Science, Atheism, Antiteism, Antireligionism, Ignosticism, Left-Libertarianism, Anarchism, Socialism, Mutualism, Axiology, Metaphysics, LGBTQI, Philosophy, Advocacy, Activism, Mental Health, Psychology, Archaeology, Social Work, Sexual Rights, Marriage Rights, Woman’s Rights, Gender Rights, Child Rights, Secular Rights, Race Equality, Ageism/Disability Equality, Etc. And a far-leftist, “Anarcho-Humanist.”
I am not a good fit in the atheist movement that is mostly pro-capitalist, I am anti-capitalist. Mostly pro-skeptic, I am a rationalist not valuing skepticism. Mostly pro-agnostic, I am anti-agnostic. Mostly limited to anti-Abrahamic religions, I am an anti-religionist.
To me, the “male god” seems to have either emerged or become prominent around 7,000 years ago, whereas the now favored monotheism “male god” is more like 4,000 years ago or so. To me, the “female goddess” seems to have either emerged or become prominent around 11,000-10,000 years ago or so, losing the majority of its once prominence around 2,000 years ago due largely to the now favored monotheism “male god” that grow in prominence after 4,000 years ago or so.
My Thought on the Evolution of Gods?
Animal protector deities from old totems/spirit animal beliefs come first to me, 13,000/12,000 years ago, then women as deities 11,000/10,000 years ago, then male gods around 7,000/8,000 years ago. Moralistic gods around 5,000/4,000 years ago, and monotheistic gods around 4,000/3,000 years ago.
To me, animal gods were likely first related to totemism animals around 13,000 to 12,000 years ago or older. Female as goddesses was next to me, 11,000 to 10,000 years ago or so with the emergence of agriculture. Then male gods come about 8,000 to 7,000 years ago with clan wars. Many monotheism-themed religions started in henotheism, emerging out of polytheism/paganism.


Damien Marie AtHope (Said as “At” “Hope”)/(Autodidact Polymath but not good at math):
Axiological Atheist, Anti-theist, Anti-religionist, Secular Humanist, Rationalist, Writer, Artist, Jeweler, Poet, “autodidact” Philosopher, schooled in Psychology, and “autodidact” Armchair Archaeology/Anthropology/Pre-Historian (Knowledgeable in the range of: 1 million to 5,000/4,000 years ago). I am an anarchist socialist politically. Reasons for or Types of Atheism
My Website, My Blog, & Short-writing or Quotes, My YouTube, Twitter: @AthopeMarie, and My Email: damien.marie.athope@gmail.com